

The absence of Top3 reveals an interaction between the Sgs1 and Pif1 DNA helicases in Saccharomyces cerevisiae
- PMID: 16816432
- PMCID: PMC1602079
- DOI: 10.1534/genetics.104.036905
The absence of Top3 reveals an interaction between the Sgs1 and Pif1 DNA helicases in Saccharomyces cerevisiae
Abstract
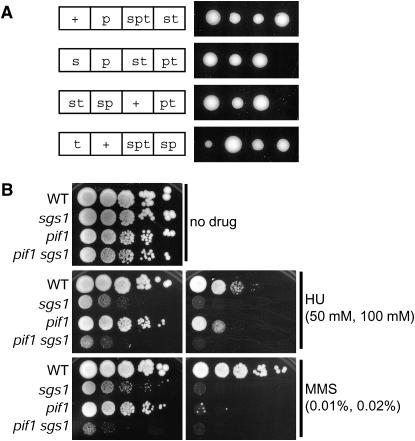
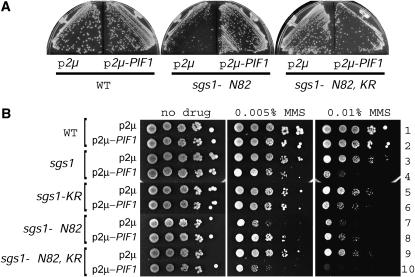
RecQ DNA helicases and Topo III topoisomerases have conserved genetic, physical, and functional interactions that are consistent with a model in which RecQ creates a recombination-dependent substrate that is resolved by Topo III. The phenotype associated with Topo III loss suggests that accumulation of a RecQ-created substrate is detrimental. In yeast, mutation of the TOP3 gene encoding Topo III causes pleiotropic defects that are suppressed by deletion of the RecQ homolog Sgs1. We searched for gene dosage suppressors of top3 and identified Pif1, a DNA helicase that acts with polarity opposite to that of Sgs1. Pif1 overexpression suppresses multiple top3 defects, but exacerbates sgs1 and sgs1 top3 defects. Furthermore, Pif1 helicase activity is essential in the absence of Top3 in an Sgs1-dependent manner. These data clearly demonstrate that Pif1 helicase activity is required to counteract Sgs1 helicase activity that has become uncoupled from Top3. Pif1 genetic interactions with the Sgs1-Top3 pathway are dependent upon homologous recombination. We also find that Pif1 is recruited to DNA repair foci and that the frequency of these foci is significantly increased in top3 mutants. Our results support a model in which Pif1 has a direct role in the prevention or repair of Sgs1-induced DNA damage that accumulates in top3 mutants.
Figures







Similar articles
-
Mutations in homologous recombination genes rescue top3 slow growth in Saccharomyces cerevisiae.Genetics. 2002 Oct;162(2):647-62. doi: 10.1093/genetics/162.2.647. Genetics. 2002. PMID: 12399378 Free PMC article.
-
Yeast Rmi1/Nce4 controls genome stability as a subunit of the Sgs1-Top3 complex.Mol Cell Biol. 2005 Jun;25(11):4476-87. doi: 10.1128/MCB.25.11.4476-4487.2005. Mol Cell Biol. 2005. PMID: 15899853 Free PMC article.
-
WRN helicase defective in the premature aging disorder Werner syndrome genetically interacts with topoisomerase 3 and restores the top3 slow growth phenotype of sgs1 top3.Aging (Albany NY). 2009 Feb 5;1(2):219-33. doi: 10.18632/aging.100020. Aging (Albany NY). 2009. PMID: 20157511 Free PMC article.
-
[Functional analysis of yeast homologue gene associated with human DNA helicase causative syndromes].Kokuritsu Iyakuhin Shokuhin Eisei Kenkyusho Hokoku. 2002;(120):53-74. Kokuritsu Iyakuhin Shokuhin Eisei Kenkyusho Hokoku. 2002. PMID: 12638184 Review. Japanese.
-
Maintenance of Yeast Genome Integrity by RecQ Family DNA Helicases.Genes (Basel). 2020 Feb 18;11(2):205. doi: 10.3390/genes11020205. Genes (Basel). 2020. PMID: 32085395 Free PMC article. Review.
Cited by
-
The Schizosaccharomyces pombe Pfh1p DNA helicase is essential for the maintenance of nuclear and mitochondrial DNA.Mol Cell Biol. 2008 Nov;28(21):6594-608. doi: 10.1128/MCB.00191-08. Epub 2008 Aug 25. Mol Cell Biol. 2008. PMID: 18725402 Free PMC article.
-
A Delicate Balance Between Repair and Replication Factors Regulates Recombination Between Divergent DNA Sequences in Saccharomyces cerevisiae.Genetics. 2016 Feb;202(2):525-40. doi: 10.1534/genetics.115.184093. Epub 2015 Dec 17. Genetics. 2016. PMID: 26680658 Free PMC article.
-
Mitochondrial helicases and mitochondrial genome maintenance.Mech Ageing Dev. 2010 Jul-Aug;131(7-8):503-10. doi: 10.1016/j.mad.2010.04.009. Epub 2010 May 31. Mech Ageing Dev. 2010. PMID: 20576512 Free PMC article.
-
Identification of Saccharomyces cerevisiae Genes Whose Deletion Causes Synthetic Effects in Cells with Reduced Levels of the Nuclear Pif1 DNA Helicase.G3 (Bethesda). 2015 Oct 19;5(12):2913-8. doi: 10.1534/g3.115.021139. G3 (Bethesda). 2015. PMID: 26483010 Free PMC article.
-
Fault tolerance in protein interaction networks: stable bipartite subgraphs and redundant pathways.PLoS One. 2009;4(4):e5364. doi: 10.1371/journal.pone.0005364. Epub 2009 Apr 28. PLoS One. 2009. PMID: 19399174 Free PMC article.
References
-
- Akada, R., J. Yamamoto and I. Yamashita, 1997. Screening and identification of yeast sequences that cause growth inhibition when overexpressed. Mol. Gen. Genet. 254: 267–274. - PubMed
-
- Bennett, R. J., J. L. Keck and J. C. Wang, 1999. Binding specificity determines polarity of DNA unwinding by the Sgs1 protein of Saccharomyces cerevisiae. J. Mol. Biol. 289: 235–248. - PubMed
-
- Bennett, R. J., M. F. Noirot-Gros and J. C. Wang, 2000. Interaction between yeast sgs1 helicase and DNA topoisomerase III. J. Biol. Chem. 275: 26898–26905. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
