

A disease- and phosphorylation-related nonmechanical function for keratin 8
- PMID: 16818723
- PMCID: PMC2064169
- DOI: 10.1083/jcb.200602146
A disease- and phosphorylation-related nonmechanical function for keratin 8
Abstract
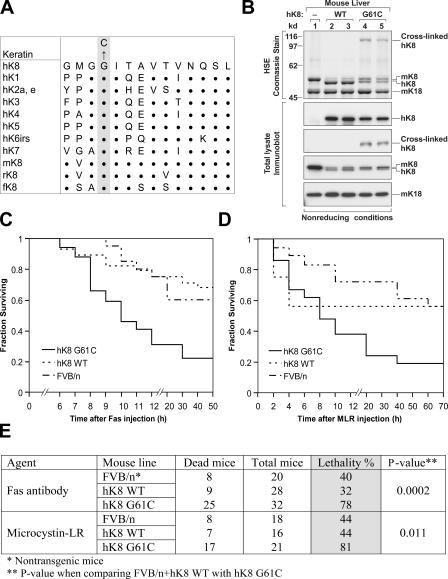
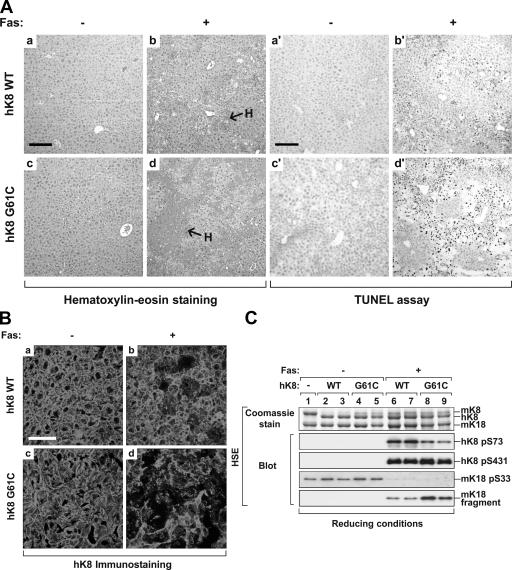
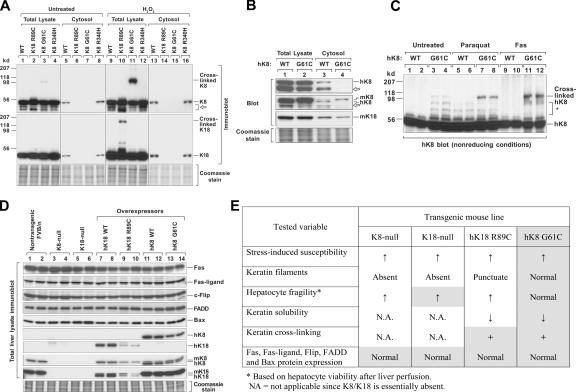
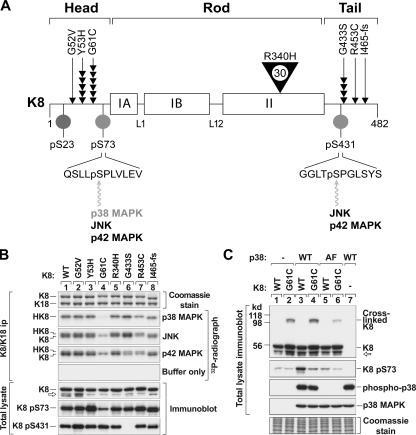
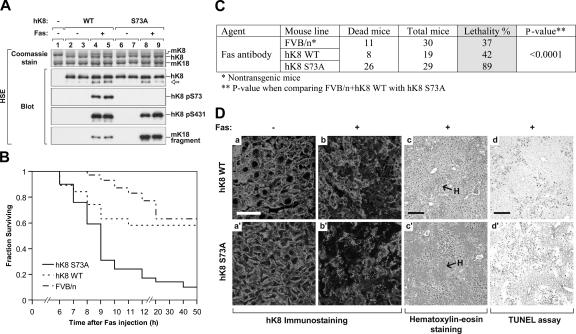
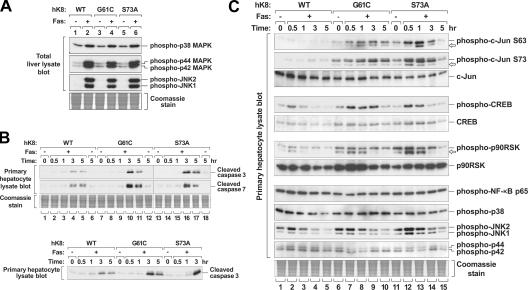
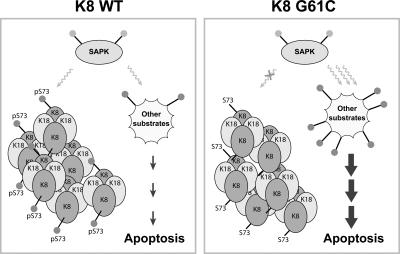
Keratin 8 (K8) variants predispose to human liver injury via poorly understood mechanisms. We generated transgenic mice that overexpress the human disease-associated K8 Gly61-to-Cys (G61C) variant and showed that G61C predisposes to liver injury and apoptosis and dramatically inhibits K8 phosphorylation at serine 73 (S73) via stress-activated kinases. This led us to generate mice that overexpress K8 S73-to-Ala (S73A), which mimicked the susceptibility of K8 G61C mice to injury, thereby providing a molecular link between K8 phosphorylation and disease-associated mutation. Upon apoptotic stimulation, G61C and S73A hepatocytes have persistent and increased nonkeratin proapoptotic substrate phosphorylation by stress-activated kinases, compared with wild-type hepatocytes, in association with an inability to phosphorylate K8 S73. Our findings provide the first direct link between patient-related human keratin variants and liver disease predisposition. The highly abundant cytoskeletal protein K8, and possibly other keratins with the conserved S73-containing phosphoepitope, can protect tissue from injury by serving as a phosphate "sponge" for stress-activated kinases and thereby provide a novel nonmechanical function for intermediate filament proteins.
Figures
Similar articles
-
Keratin mutation in transgenic mice predisposes to Fas but not TNF-induced apoptosis and massive liver injury.Hepatology. 2003 May;37(5):1006-14. doi: 10.1053/jhep.2003.50181. Hepatology. 2003. PMID: 12717381
-
Keratins let liver live: Mutations predispose to liver disease and crosslinking generates Mallory-Denk bodies.Hepatology. 2007 Nov;46(5):1639-49. doi: 10.1002/hep.21976. Hepatology. 2007. PMID: 17969036 Review.
-
Protein phosphatase inhibition in normal and keratin 8/18 assembly-incompetent mouse strains supports a functional role of keratin intermediate filaments in preserving hepatocyte integrity.Hepatology. 1998 Jul;28(1):116-28. doi: 10.1002/hep.510280117. Hepatology. 1998. PMID: 9657104
-
Novel insights into changes in biochemical properties of keratins 8 and 18 in griseofulvin-induced toxic liver injury.Exp Mol Pathol. 2010 Oct;89(2):117-25. doi: 10.1016/j.yexmp.2010.07.004. Epub 2010 Jul 17. Exp Mol Pathol. 2010. PMID: 20643122
-
Keratin modifications and solubility properties in epithelial cells and in vitro.Subcell Biochem. 1998;31:105-40. Subcell Biochem. 1998. PMID: 9932491 Review.
Cited by
-
Post-translational modifications of intermediate filament proteins: mechanisms and functions.Nat Rev Mol Cell Biol. 2014 Mar;15(3):163-77. doi: 10.1038/nrm3753. Nat Rev Mol Cell Biol. 2014. PMID: 24556839 Free PMC article. Review.
-
Keratin mutation predisposes to mouse liver fibrosis and unmasks differential effects of the carbon tetrachloride and thioacetamide models.Gastroenterology. 2008 Apr;134(4):1169-79. doi: 10.1053/j.gastro.2008.01.035. Epub 2008 Jan 18. Gastroenterology. 2008. PMID: 18395095 Free PMC article.
-
"Panta rhei": Perpetual cycling of the keratin cytoskeleton.Bioarchitecture. 2011 Jan;1(1):39-44. doi: 10.4161/bioa.1.1.14815. Bioarchitecture. 2011. PMID: 21866261 Free PMC article.
-
Absence of keratin 8 confers a paradoxical microflora-dependent resistance to apoptosis in the colon.Proc Natl Acad Sci U S A. 2011 Jan 25;108(4):1445-50. doi: 10.1073/pnas.1010833108. Epub 2011 Jan 10. Proc Natl Acad Sci U S A. 2011. PMID: 21220329 Free PMC article.
-
The human keratins: biology and pathology.Histochem Cell Biol. 2008 Jun;129(6):705-33. doi: 10.1007/s00418-008-0435-6. Epub 2008 May 7. Histochem Cell Biol. 2008. PMID: 18461349 Free PMC article. Review.
References
-
- Baines, C.P., and J.D. Molkentin. 2005. STRESS signaling pathways that modulate cardiac myocyte apoptosis. J. Mol. Cell. Cardiol. 38:47–62. - PubMed
-
- Baribault, H., J. Penner, R.V. Iozzo, and M. Wilson-Heiner. 1994. Colorectal hyperplasia and inflammation in keratin 8-deficient FVB/N mice. Genes Dev. 8:2964–2973. - PubMed
-
- Behrens, A., M. Sibilia, and E.F. Wagner. 1999. Amino-terminal phosphorylation of c-Jun regulates stress-induced apoptosis and cellular proliferation. Nat. Genet. 21:326–329. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
