

Inhibition of hemoglobin S polymerization in vitro by a novel 15-mer EF-helix beta73 histidine-containing peptide
- PMID: 16819835
- PMCID: PMC2593912
- DOI: 10.1021/bi0604734
Inhibition of hemoglobin S polymerization in vitro by a novel 15-mer EF-helix beta73 histidine-containing peptide
Abstract
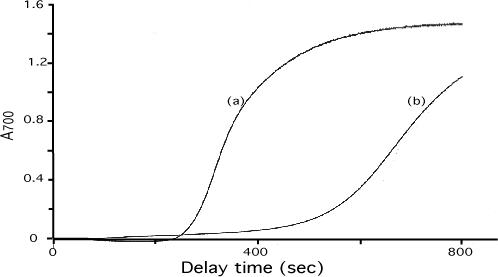
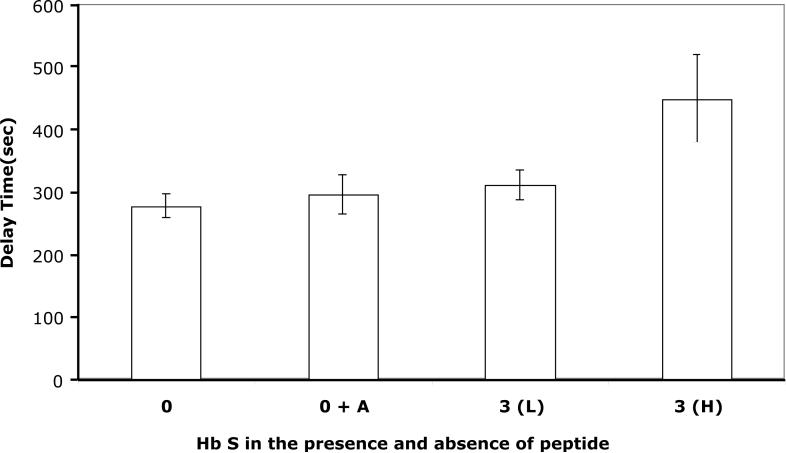
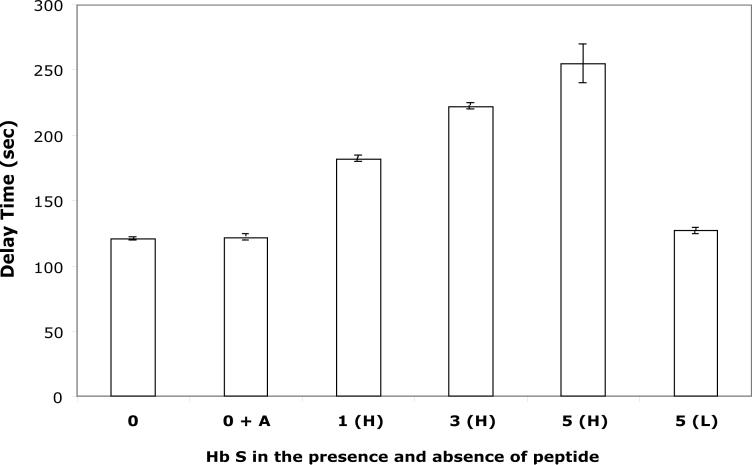
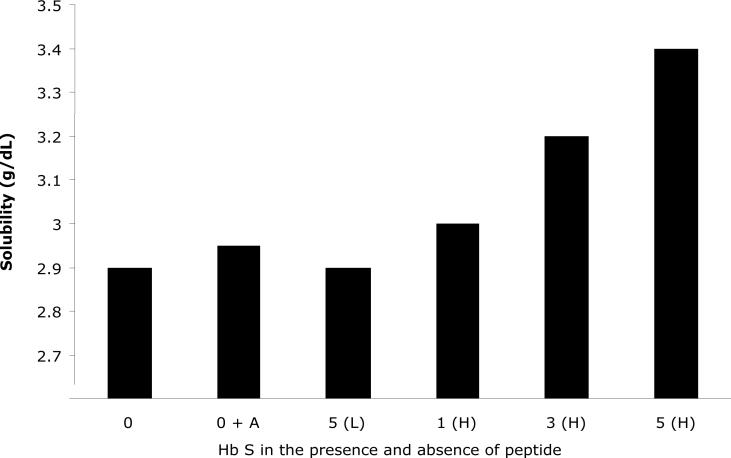
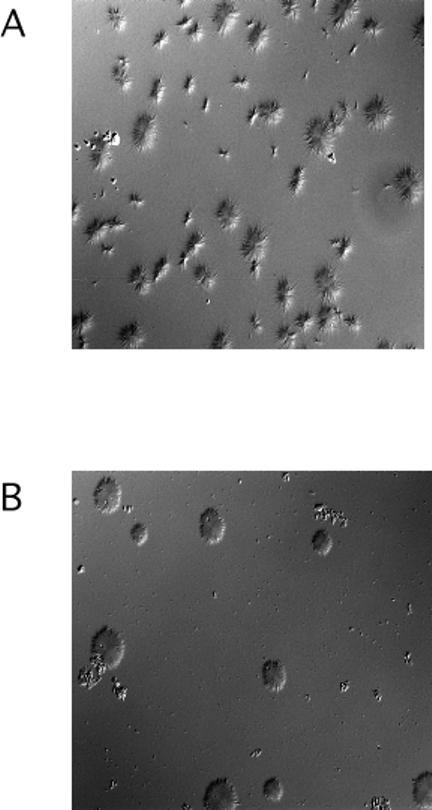
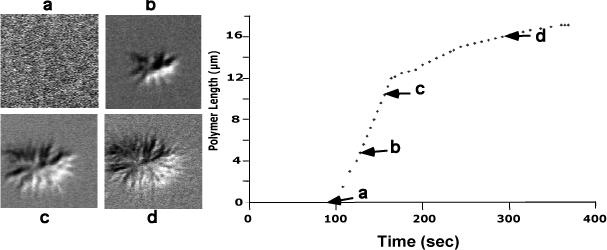
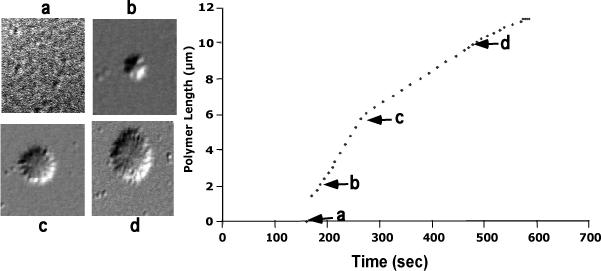
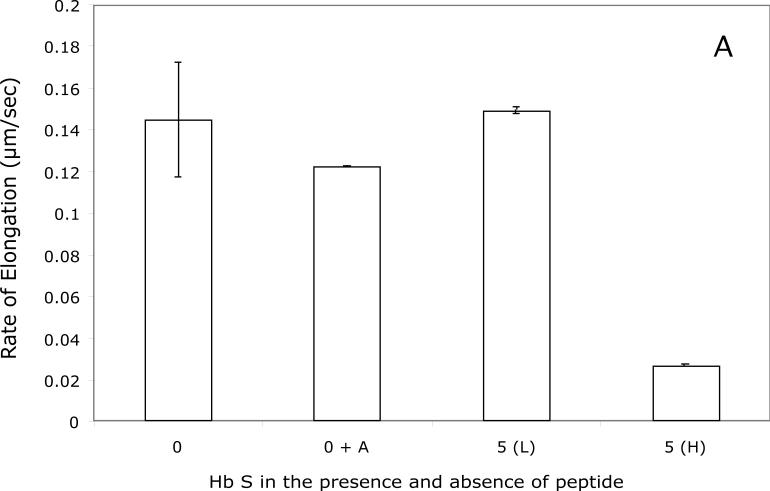
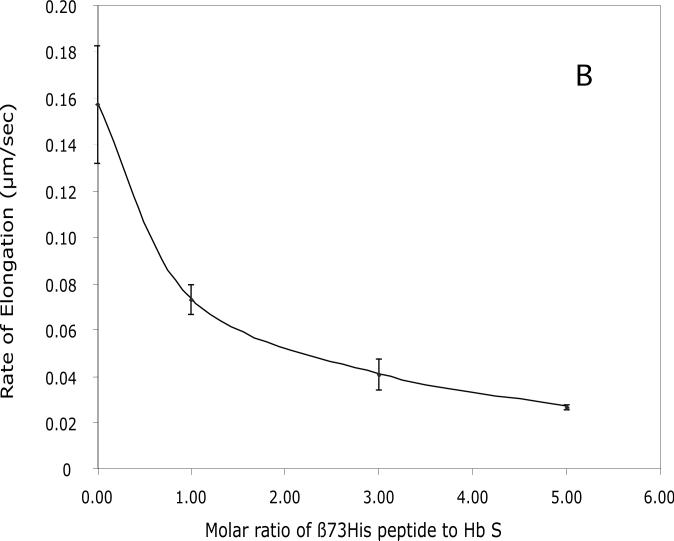
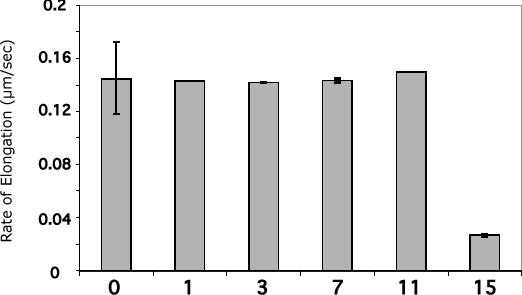
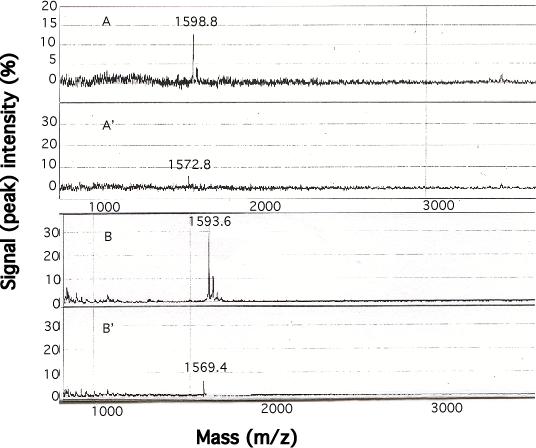
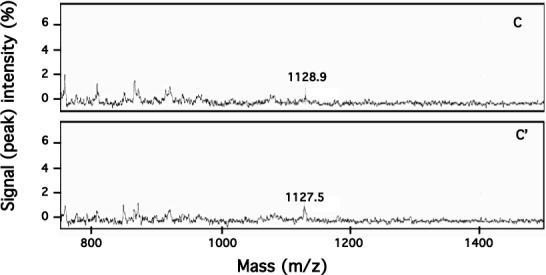
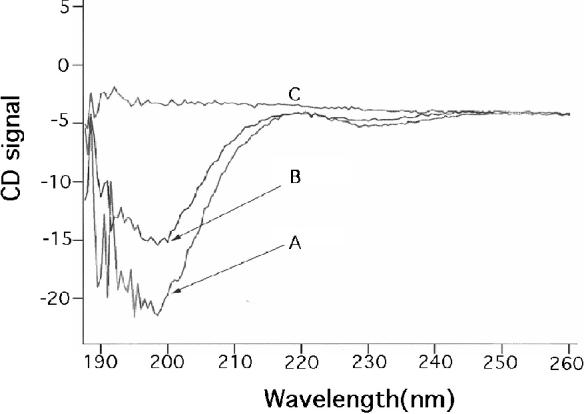
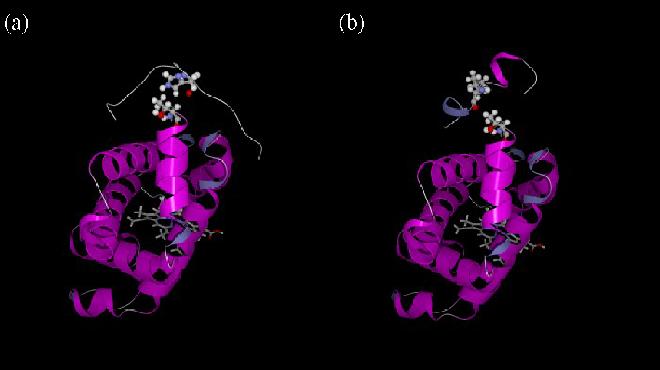
Our mutational studies on Hb S showed that the Hb S beta73His variant (beta6Val and beta73His) promoted polymerization, while Hb S beta73Leu (beta6Val and beta73Leu) inhibited polymerization. On the basis of these results, we speculated that EF-helix peptides containing beta73His interact with beta4Thr in Hb S and compete with Hb S, resulting in inhibition of Hb S polymerization. We, therefore, studied inhibitory effects of 15-, 11-, 7-, and 3-mer EF-helix peptides containing beta73His on Hb S polymerization. The delay time prior to Hb S polymerization increased only in the presence of the 15-mer His peptide; the higher the amount, the longer the delay time. DIC image analysis also showed that the fiber elongation rate for Hb S polymers decreased with increasing concentration of the 15-mer His peptide. In contrast, the same 15-mer peptide containing beta73Leu instead of His and peptides shorter than 11 amino acids containing beta73His including His alone showed little effect on the kinetics of polymerization and elongation of polymers. Analysis by protein-chip arrays showed that only the 15-mer beta73His peptide interacted with Hb S. CD spectra of the 15-mer beta73His peptide did not show a specific helical structure; however, computer docking analysis suggested a lower energy for interaction of Hb S with the 15-mer beta73His peptide compared to peptides containing other amino acids at this position. These results suggest that the 15-mer beta73His peptide interacts with Hb S via the beta4Thr in the betaS-globin chain in Hb S. This interaction may influence hydrogen bond interaction between beta73Asp and beta4Thr in Hb S polymers and interfere in hydrophobic interactions of beta6Val, leading to inhibition of Hb S polymerization.
Figures
Similar articles
-
Effects of different beta73 amino acids on formation of 14-stranded fibers of Hb S versus double-stranded crystals of Hb C-Harlem.Biochemistry. 2003 Apr 22;42(15):4476-84. doi: 10.1021/bi026740x. Biochemistry. 2003. PMID: 12693943
-
Alizarin interaction with sickle hemoglobin: elucidation of their anti-sickling properties by multi-spectroscopic and molecular modeling techniques.J Biomol Struct Dyn. 2019 Oct;37(17):4614-4631. doi: 10.1080/07391102.2018.1557557. Epub 2019 Jan 24. J Biomol Struct Dyn. 2019. PMID: 30558488
-
Polymerization of recombinant hemoglobin F gamma E6V and hemoglobin F gamma E6V, gamma Q87T alone, and in mixtures with hemoglobin S.Blood. 1996 Feb 15;87(4):1617-24. Blood. 1996. PMID: 8608256
-
The Hb A variant (beta73 Asp-->Leu) disrupts Hb S polymerization by a novel mechanism.J Mol Biol. 2006 Sep 22;362(3):528-38. doi: 10.1016/j.jmb.2006.07.047. Epub 2006 Jul 28. J Mol Biol. 2006. PMID: 16926024
-
Role of the beta4Thr-beta73Asp hydrogen bond in HbS polymer and domain formation from multinucleate-containing clusters.Biochemistry. 2008 May 13;47(19):5441-9. doi: 10.1021/bi800149u. Epub 2008 Apr 18. Biochemistry. 2008. PMID: 18419131
Cited by
-
Rational Drug Design of Peptide-Based Therapies for Sickle Cell Disease.Molecules. 2019 Dec 12;24(24):4551. doi: 10.3390/molecules24244551. Molecules. 2019. PMID: 31842406 Free PMC article. Review.
-
Relationship between beta4 hydrogen bond and beta6 hydrophobic interactions during aggregate, fiber or crystal formation in oversaturated solutions of hemoglobin A and S.Arch Biochem Biophys. 2009 Jan 15;481(2):137-44. doi: 10.1016/j.abb.2008.11.006. Epub 2008 Nov 13. Arch Biochem Biophys. 2009. PMID: 19022217 Free PMC article.
References
-
- Eaton WA, Hofrichter J. Sickle cell hemoglobin polymerization. Adv Protein Chem. 1990;40:63–279. - PubMed
-
- Bunn HF, Forget BG. Hemoglobin: Molecular and Clinical Aspects. Sanders; Philadelphia.: 1996.
-
- Ferrone FA, Hofrichter J, Eaton WA. Kinetics of sickle hemoglobin polymerization. II. A double nucleation mechanism. J Mol Biol. 1985;183:611–631. - PubMed
-
- Embury SH, Hebbel RP, Mohandus N, Steinberg MH. SICKLE CELL DISEASE: Basic principles and clinical practice. Raven Press; New York: 1994.
-
- Harrington DJ, Adachi K, Royer WE., Jr. The high resolution crystal structure of deoxyhemoglobin S. J Mol Biol. 1997;272:398–407. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
