

Alteration of the rugose phenotype in waaG and ddhC mutants of Salmonella enterica serovar Typhimurium DT104 is associated with inverse production of curli and cellulose
- PMID: 16820499
- PMCID: PMC1489332
- DOI: 10.1128/AEM.02868-05
Alteration of the rugose phenotype in waaG and ddhC mutants of Salmonella enterica serovar Typhimurium DT104 is associated with inverse production of curli and cellulose
Abstract
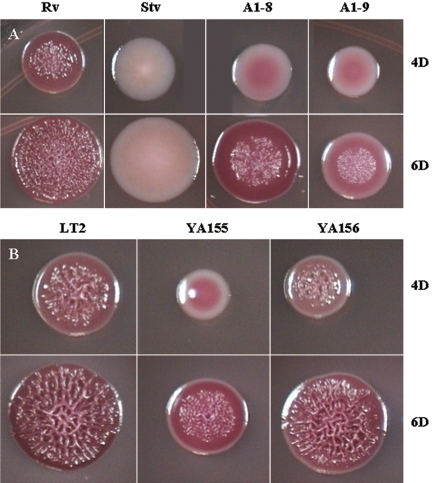
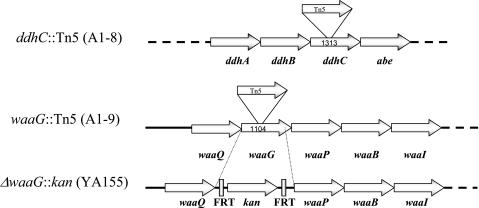
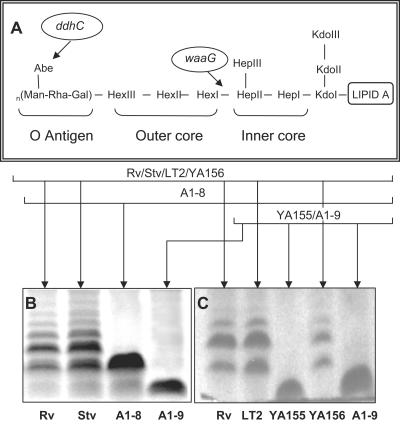
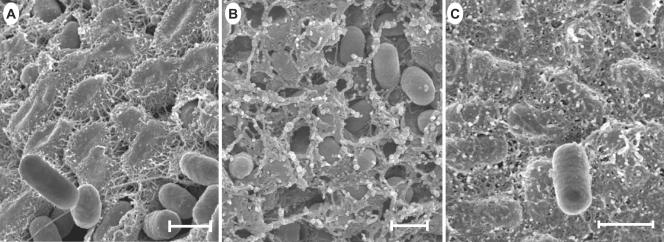
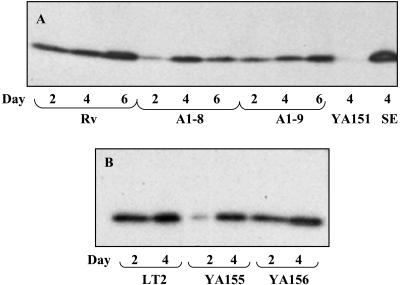
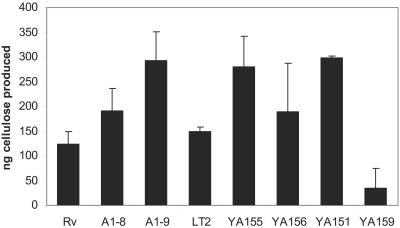
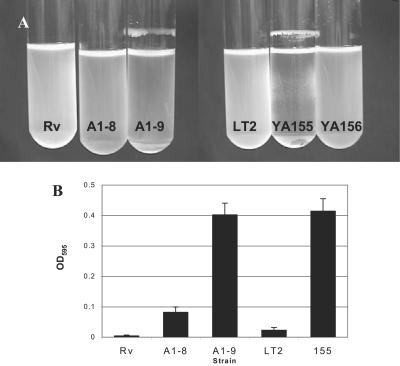
The rugose (also known as wrinkled or rdar) phenotype in Salmonella enterica serovar Typhimurium DT104 Rv has been associated with cell aggregation and the ability, at low temperature under low-osmolarity conditions, to form pellicles and biofilms. Two Tn5 insertion mutations in genes that are involved in lipopolysaccharide (LPS) synthesis, ddhC (A1-8) and waaG (A1-9), of Rv resulted in diminished expression of colony rugosity. Scanning electron micrographs revealed that the ddhC mutant showed reduced amounts of extracellular matrix, while there was relatively more, profuse matrix production in the waaG mutant, compared to Rv. Both mutants appeared to produce decreased levels of curli, as judged by Western blot assays probed with anti-AgfA (curli) antibodies but, surprisingly, were observed to have increased amounts of cellulose relative to Rv. Comparison with a non-curli-producing mutant suggested that the alteration in curli production may have engendered the increased presence of cellulose. While both mutants had impaired biofilm formation when grown in rich medium with low osmolarity, they constitutively formed larger amounts of biofilms when the growth medium was supplemented with either glucose or a combination of glucose and NaCl. These observations indicated that LPS alterations may have opposing effects on biofilm formation in these mutants, depending upon either the presence or the absence of these osmolytes. The phenotypes of the waaG mutant were further confirmed in a constructed, nonpolar deletion mutant of S. enterica serovar Typhimurium LT2, where restoration to the wild-type phenotypes was accomplished by complementation. These results highlight the importance of an integral LPS, at both the O-antigen and core polysaccharide levels, in the modulation of curli protein and cellulose production, as well as in biofilm formation, thereby adding another potential component to the complex regulatory system which governs multicellular behaviors in S. enterica serovar Typhimurium.
Figures
Similar articles
-
Role of the GGDEF protein family in Salmonella cellulose biosynthesis and biofilm formation.Mol Microbiol. 2004 Oct;54(1):264-77. doi: 10.1111/j.1365-2958.2004.04269.x. Mol Microbiol. 2004. PMID: 15458421
-
Biofilm formation in field strains of Salmonella enterica serovar Typhimurium: identification of a new colony morphology type and the role of SGI1 in biofilm formation.Vet Microbiol. 2008 Jun 22;129(3-4):360-6. doi: 10.1016/j.vetmic.2007.12.006. Epub 2007 Dec 23. Vet Microbiol. 2008. PMID: 18242887
-
Analyses of the red-dry-rough phenotype of an Escherichia coli O157:H7 strain and its role in biofilm formation and resistance to antibacterial agents.Appl Environ Microbiol. 2006 Apr;72(4):2564-72. doi: 10.1128/AEM.72.4.2564-2572.2006. Appl Environ Microbiol. 2006. PMID: 16597958 Free PMC article.
-
The csgD promoter, a control unit for biofilm formation in Salmonella typhimurium.Res Microbiol. 2003 Dec;154(10):659-67. doi: 10.1016/j.resmic.2003.08.005. Res Microbiol. 2003. PMID: 14643403 Review.
-
Regulation of biofilm formation in Salmonella enterica serovar Typhimurium.Future Microbiol. 2014;9(11):1261-82. doi: 10.2217/fmb.14.88. Future Microbiol. 2014. PMID: 25437188 Review.
Cited by
-
Cronobacter: an emergent pathogen causing meningitis to neonates through their feeds.Sci Prog. 2014;97(Pt 2):154-72. doi: 10.3184/003685014X13994743930498. Sci Prog. 2014. PMID: 25108996 Free PMC article.
-
The gpsX gene encoding a glycosyltransferase is important for polysaccharide production and required for full virulence in Xanthomonas citri subsp. citri.BMC Microbiol. 2012 Mar 9;12:31. doi: 10.1186/1471-2180-12-31. BMC Microbiol. 2012. PMID: 22404966 Free PMC article.
-
Dimethyl adenosine transferase (KsgA) contributes to cell-envelope fitness in Salmonella Enteritidis.Microbiol Res. 2018 Nov;216:108-119. doi: 10.1016/j.micres.2018.08.009. Epub 2018 Aug 23. Microbiol Res. 2018. PMID: 30269850 Free PMC article.
-
Roles of the spiA gene from Salmonella enteritidis in biofilm formation and virulence.Microbiology (Reading). 2011 Jun;157(Pt 6):1798-1805. doi: 10.1099/mic.0.046185-0. Epub 2011 Mar 17. Microbiology (Reading). 2011. PMID: 21415117 Free PMC article.
-
Only time will tell: lipopolysaccharide glycoform and biofilm-formation kinetics in Salmonella species and Escherichia coli.J Bacteriol. 2024 Oct 24;206(10):e0031824. doi: 10.1128/jb.00318-24. Epub 2024 Sep 24. J Bacteriol. 2024. PMID: 39315775 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
