

The plug domain of yeast Sec61p is important for efficient protein translocation, but is not essential for cell viability
- PMID: 16822836
- PMCID: PMC1556385
- DOI: 10.1091/mbc.e06-03-0200
The plug domain of yeast Sec61p is important for efficient protein translocation, but is not essential for cell viability
Abstract
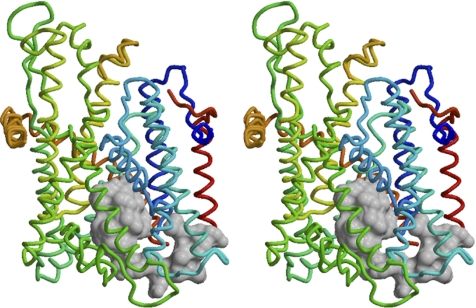
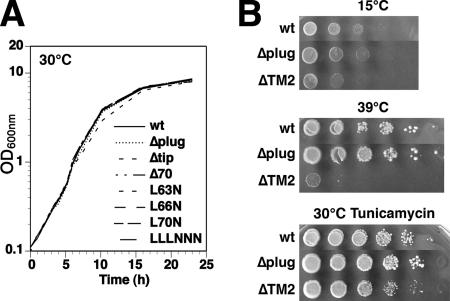
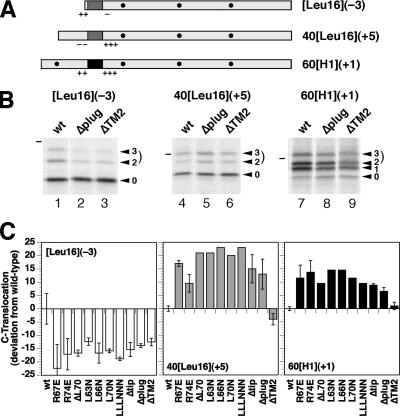
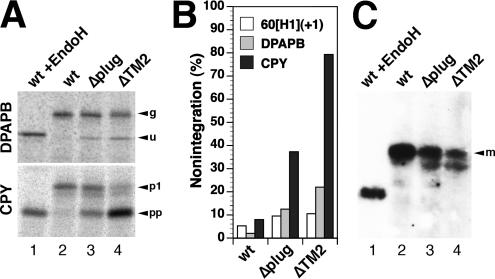
The Sec61/SecY translocon mediates translocation of proteins across the membrane and integration of membrane proteins into the lipid bilayer. The structure of the translocon revealed a plug domain blocking the pore on the lumenal side. It was proposed to be important for gating the protein conducting channel and for maintaining the permeability barrier in its unoccupied state. Here, we analyzed in yeast the effect of introducing destabilizing point mutations in the plug domain or of its partial or complete deletion. Unexpectedly, even when the entire plug domain was deleted, cells were viable without growth phenotype. They showed an effect on signal sequence orientation of diagnostic signal-anchor proteins, a minor defect in cotranslational and a significant deficiency in posttranslational translocation. Steady-state levels of the mutant protein were reduced, and when coexpressed with wild-type Sec61p, the mutant lacking the plug competed poorly for complex partners. The results suggest that the plug is unlikely to be important for sealing the translocation pore in yeast but that it plays a role in stabilizing Sec61p during translocon formation.
Figures

References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
