

Prostasin regulates epithelial monolayer function: cell-specific Gpld1-mediated secretion and functional role for GPI anchor
- PMID: 16822939
- PMCID: PMC2271112
- DOI: 10.1152/ajpcell.00637.2005
Prostasin regulates epithelial monolayer function: cell-specific Gpld1-mediated secretion and functional role for GPI anchor
Abstract
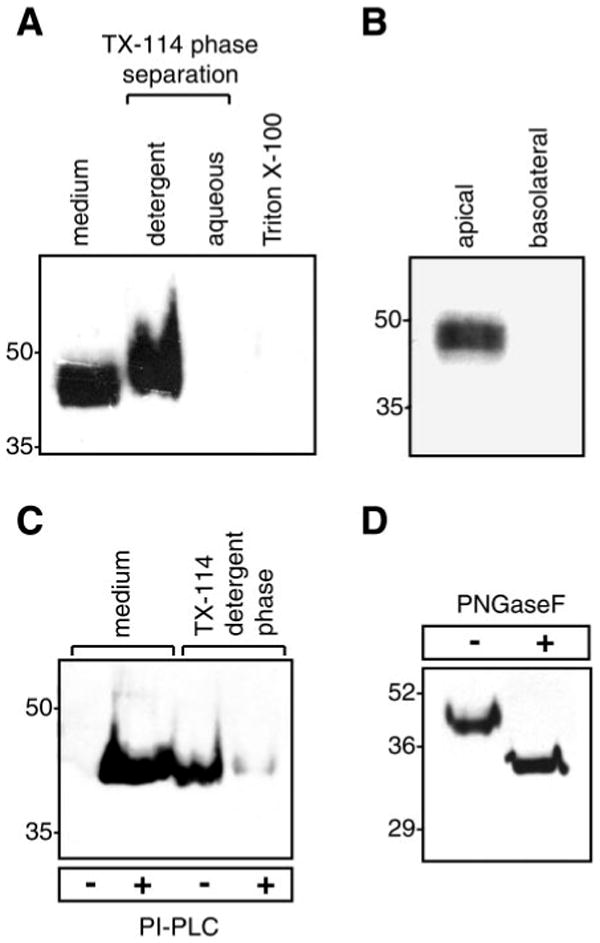
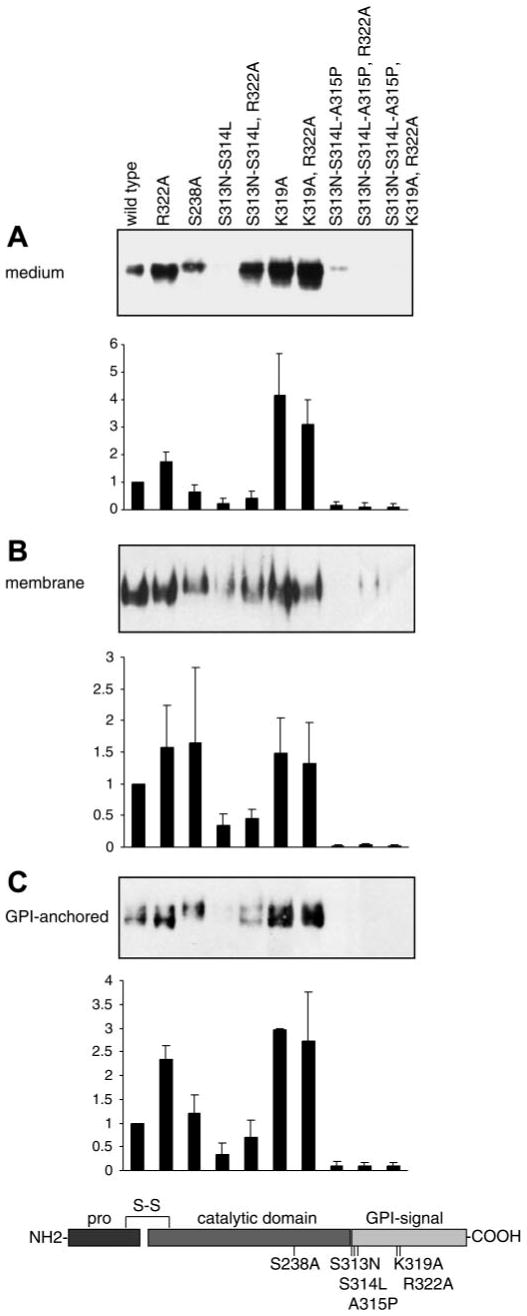
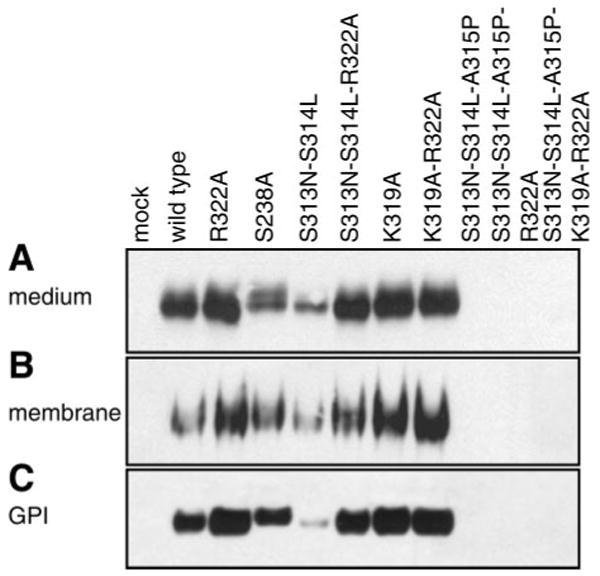
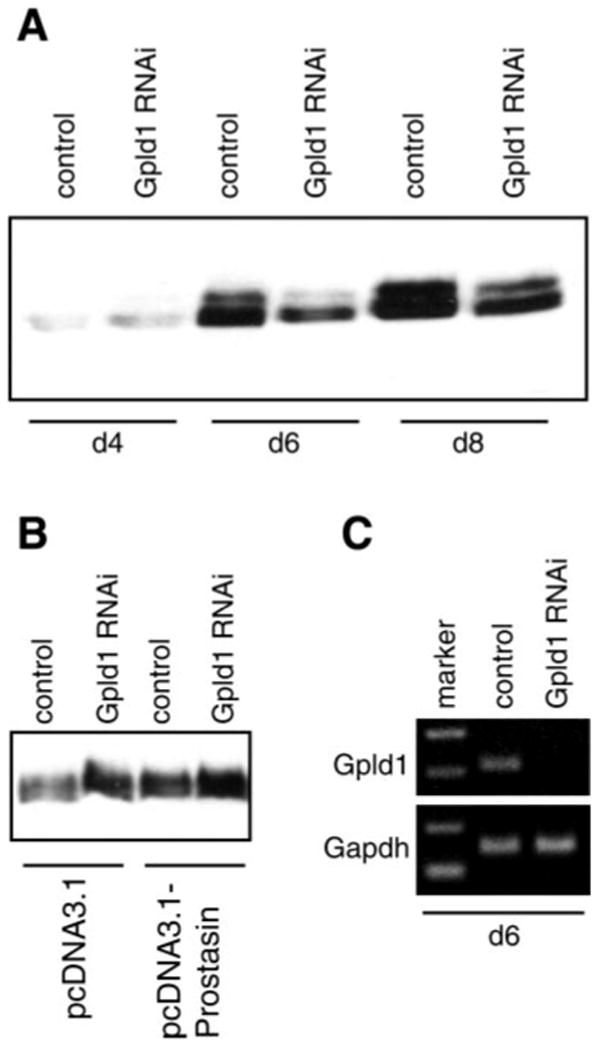
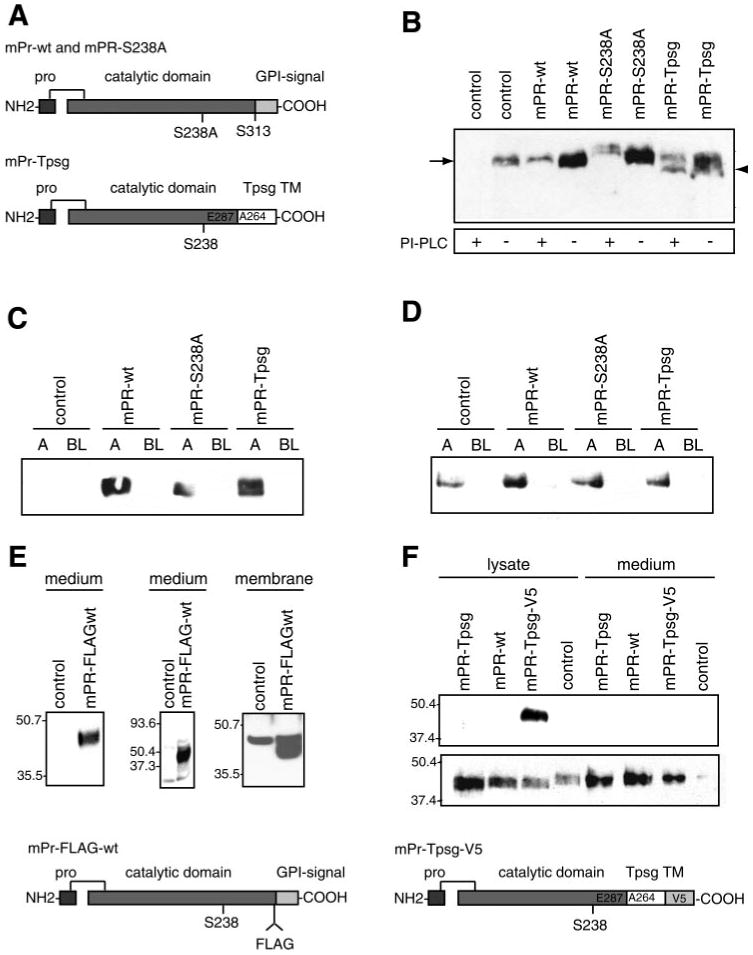
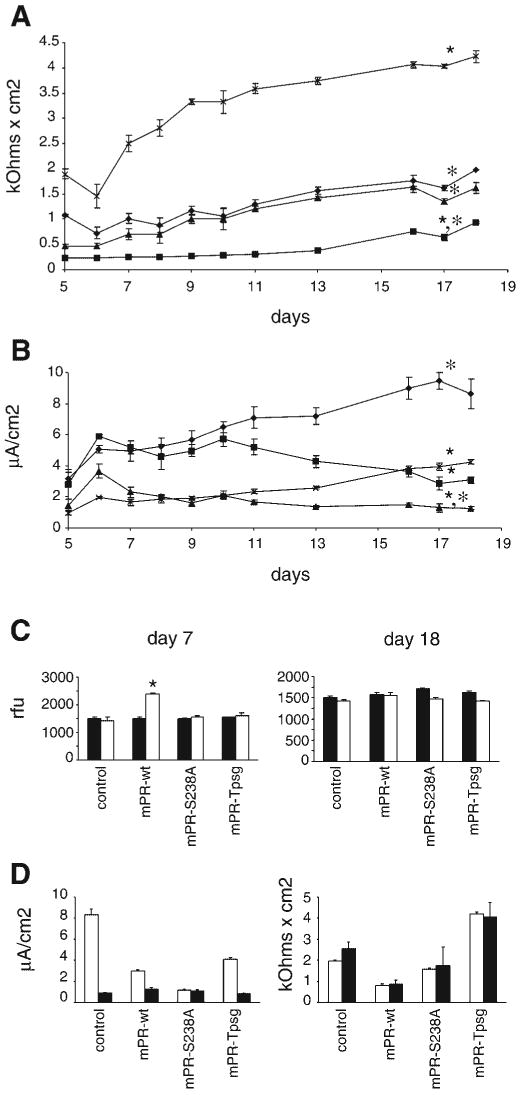
Prostasin, a trypsinlike serine peptidase, is highly expressed in prostate, kidney, and lung epithelia, where it is bound to the cell surface, secreted, or both. Prostasin activates the epithelial sodium channel (ENaC) and suppresses invasion of prostate and breast cancer cells. The studies reported here establish mechanisms of membrane anchoring and secretion in kidney and lung epithelial cells and demonstrate a critical role for prostasin in regulating epithelial monolayer function. We report that endogenous mouse prostasin is glycosylphosphatidylinositol (GPI) anchored to the cell surface and is constitutively secreted from the apical surface of kidney cortical collecting duct cells. Using site-directed mutagenesis, detergent phase separation, and RNA interference approaches, we show that prostasin secretion depends on GPI anchor cleavage by endogenous GPI-specific phospholipase D1 (Gpld1). Secretion of prostasin by kidney and lung epithelial cells, in contrast to prostate epithelium, does not depend on COOH-terminal processing at conserved Arg(322). Using stably transfected M-1 cells expressing wild-type, catalytically inactive, or chimeric transmembrane (not GPI)-anchored prostasins we establish that prostasin regulates transepithelial resistance, current, and paracellular permeability by GPI anchor- and protease activity-dependent mechanisms. These studies demonstrate a novel role for prostasin in regulating epithelial monolayer resistance and permeability in kidney epithelial cells and, furthermore, show specifically that prostasin is a critical regulator of transepithelial ion transport in M-1 cells. These functions depend on the GPI anchor as well as the peptidase activity of prostasin. These studies suggest that cell-specific Gpld1- or peptidase-dependent pathways for prostasin secretion may control prostasin functions in a tissue-specific manner.
Figures
Similar articles
-
Prostasin, a membrane-anchored serine peptidase, regulates sodium currents in JME/CF15 cells, a cystic fibrosis airway epithelial cell line.Am J Physiol Lung Cell Mol Physiol. 2004 Nov;287(5):L928-35. doi: 10.1152/ajplung.00160.2004. Epub 2004 Jul 9. Am J Physiol Lung Cell Mol Physiol. 2004. PMID: 15246975
-
Prostasin-dependent activation of epithelial Na+ channels by low plasmin concentrations.Am J Physiol Regul Integr Comp Physiol. 2009 Dec;297(6):R1733-41. doi: 10.1152/ajpregu.00321.2009. Epub 2009 Sep 30. Am J Physiol Regul Integr Comp Physiol. 2009. PMID: 19793956
-
Prostasin regulates iNOS and cyclin D1 expression by modulating protease-activated receptor-2 signaling in prostate epithelial cells.Prostate. 2009 Dec 1;69(16):1790-801. doi: 10.1002/pros.21030. Prostate. 2009. PMID: 19670249
-
The matriptase-prostasin proteolytic cascade in epithelial development and pathology.Cell Tissue Res. 2013 Feb;351(2):245-53. doi: 10.1007/s00441-012-1348-1. Epub 2012 Feb 18. Cell Tissue Res. 2013. PMID: 22350849 Review.
-
Detergent-resistant membrane microdomains and apical sorting of GPI-anchored proteins in polarized epithelial cells.Int J Med Microbiol. 2002 Feb;291(6-7):439-45. doi: 10.1078/1438-4221-00151. Int J Med Microbiol. 2002. PMID: 11890542 Review.
Cited by
-
GPI-AP release in cellular, developmental, and reproductive biology.J Lipid Res. 2016 Apr;57(4):538-45. doi: 10.1194/jlr.R063032. Epub 2015 Nov 22. J Lipid Res. 2016. PMID: 26593072 Free PMC article. Review.
-
Activity and inhibition of prostasin and matriptase on apical and basolateral surfaces of human airway epithelial cells.Am J Physiol Lung Cell Mol Physiol. 2012 Jul;303(2):L97-106. doi: 10.1152/ajplung.00303.2011. Epub 2012 May 11. Am J Physiol Lung Cell Mol Physiol. 2012. PMID: 22582115 Free PMC article.
-
Expression of prostasin and its inhibitors during colorectal cancer carcinogenesis.BMC Cancer. 2009 Jun 25;9:201. doi: 10.1186/1471-2407-9-201. BMC Cancer. 2009. PMID: 19555470 Free PMC article.
-
Mast cell tryptases and chymases in inflammation and host defense.Immunol Rev. 2007 Jun;217:141-54. doi: 10.1111/j.1600-065X.2007.00509.x. Immunol Rev. 2007. PMID: 17498057 Free PMC article. Review.
-
Prostasin expression is regulated by airway surface liquid volume and is increased in cystic fibrosis.Am J Physiol Lung Cell Mol Physiol. 2008 May;294(5):L932-41. doi: 10.1152/ajplung.00437.2007. Epub 2008 Feb 29. Am J Physiol Lung Cell Mol Physiol. 2008. PMID: 18310226 Free PMC article.
References
-
- Bauvois B. Transmembrane proteases in cell growth and invasion: new contributors to angiogenesis? Oncogene. 2004;23:317–329. - PubMed
-
- Bhagwandin V, Hau L, Clair JMS, Wolters P, Caughey G. Structure and activity of human pancreasin, a novel tryptic serine peptidase expressed primarily by the pancreas. J Biol Chem. 2003;278:3363–3371. - PubMed
-
- Brown DA, Rose JK. Sorting of GPI-anchored proteins to glycolipid-enriched membrane subdomains during transport to the apical cell surface. Cell. 1992;68:533–544. - PubMed
-
- Caughey GH, Raymond WW, Blount JL, Hau LW, Pallaoro M, Wolters PJ, Verghese GM. Characterization of human gamma-tryptases, novel members of the chromosome 16p mast cell tryptase and prostasin gene families. J Immunol. 2000;164:6566–6575. - PubMed
-
- Chen LM, Chai KX. Prostasin serine protease inhibits breast cancer invasiveness and is transcriptionally regulated by promoter DNA methylation. Int J Cancer. 2002;97:323–329. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous