

Representation of future and previous spatial goals by separate neural populations in prefrontal cortex
- PMID: 16822988
- PMCID: PMC1501084
- DOI: 10.1523/JNEUROSCI.0699-06.2006
Representation of future and previous spatial goals by separate neural populations in prefrontal cortex
Abstract
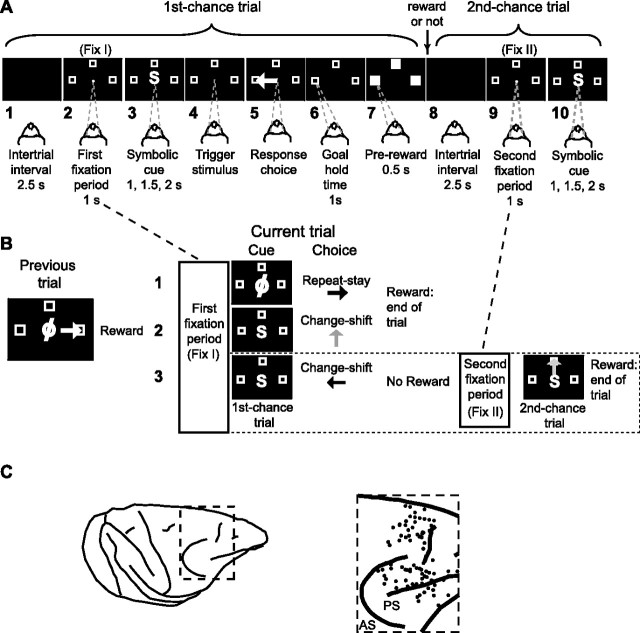
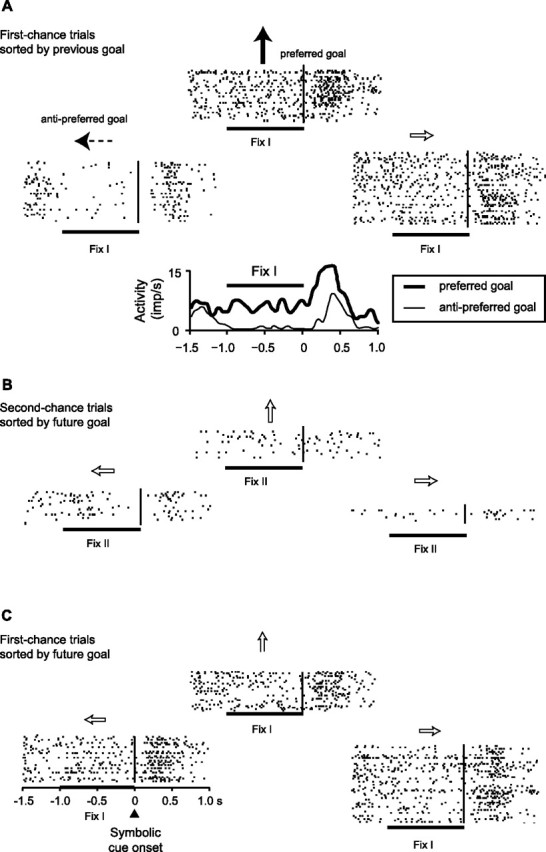
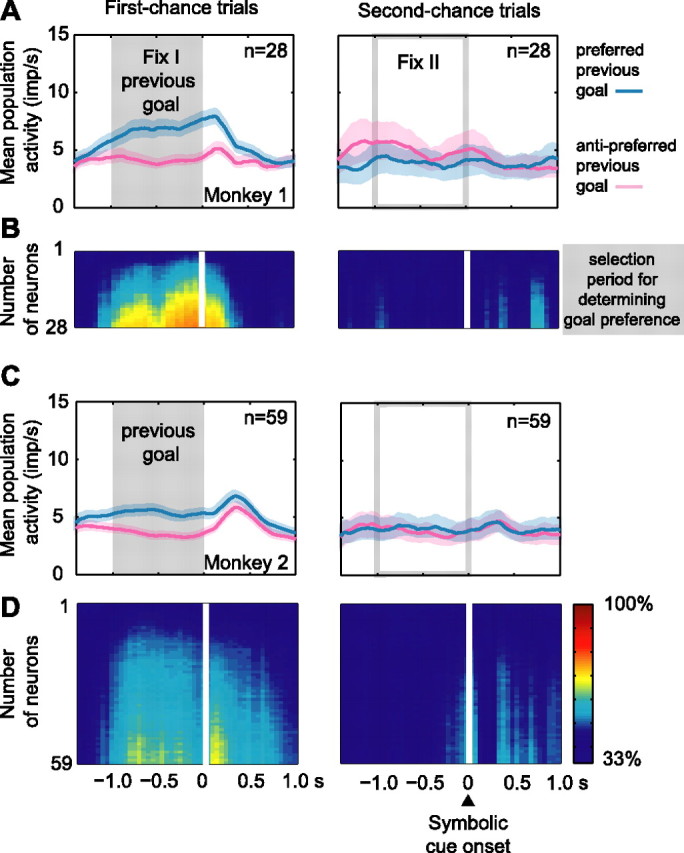
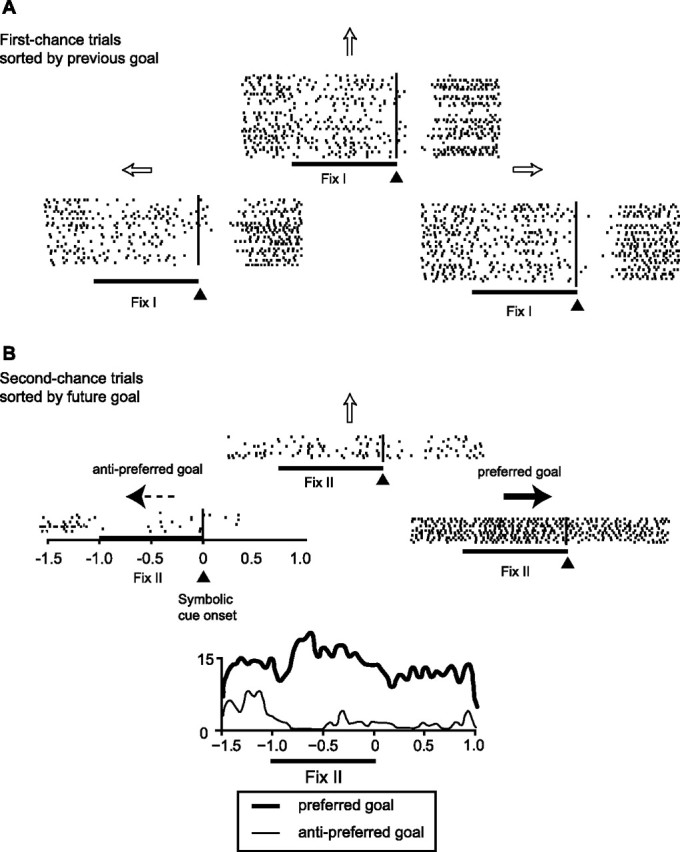
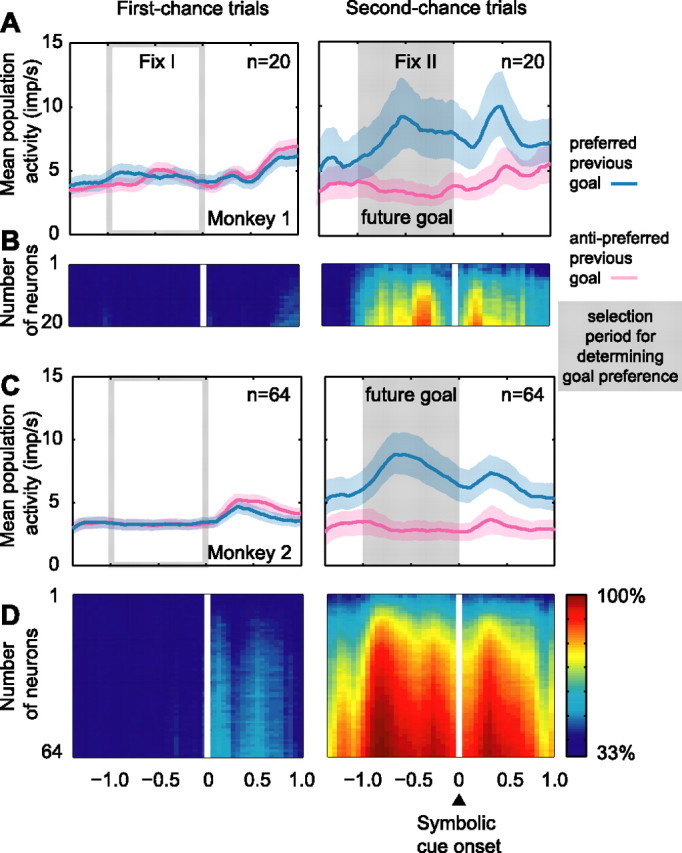
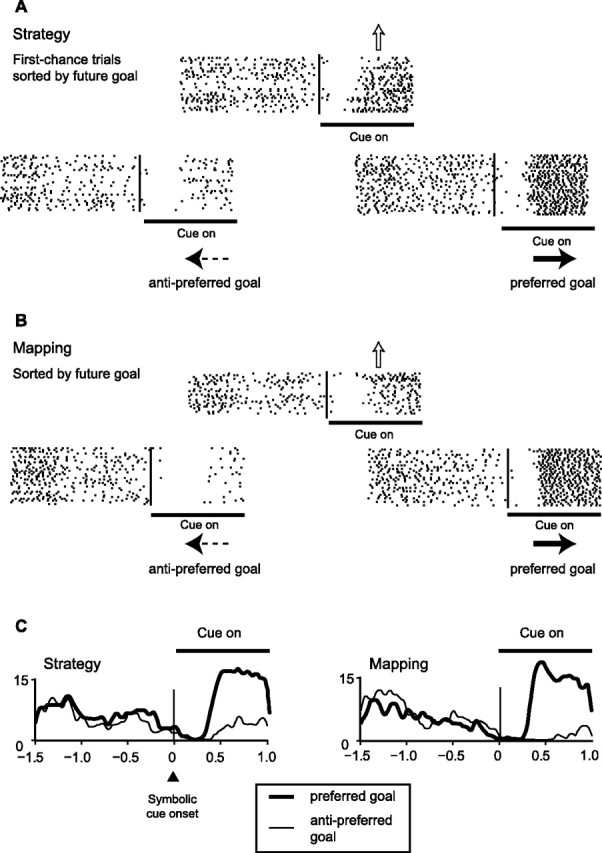
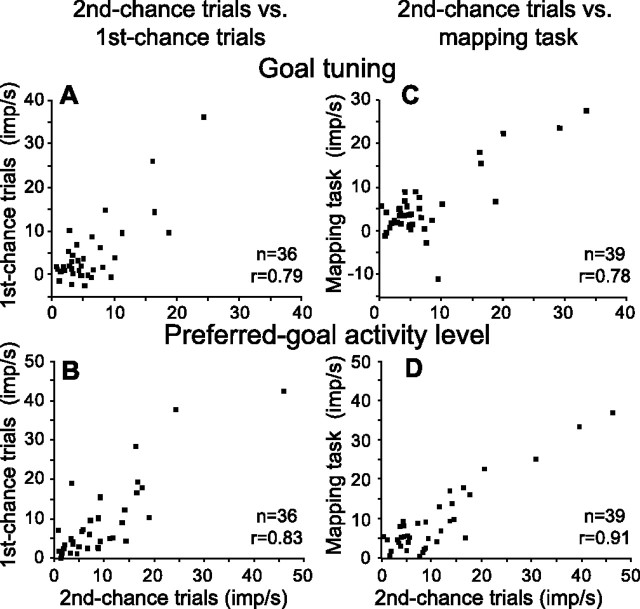
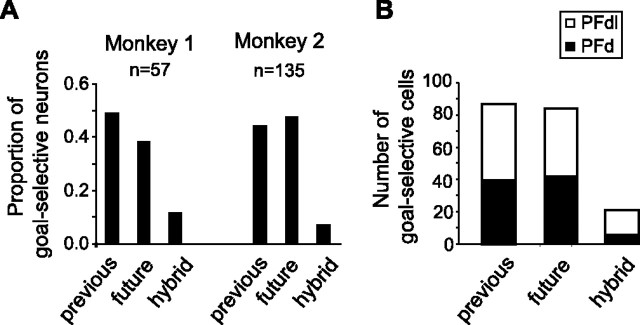
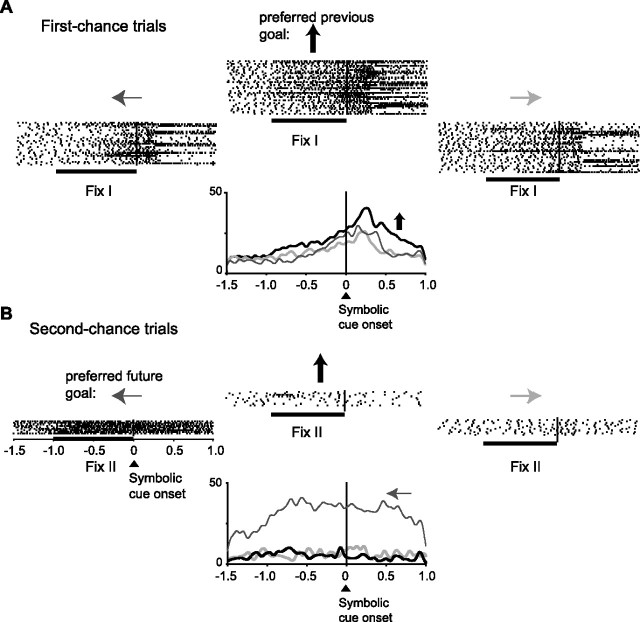
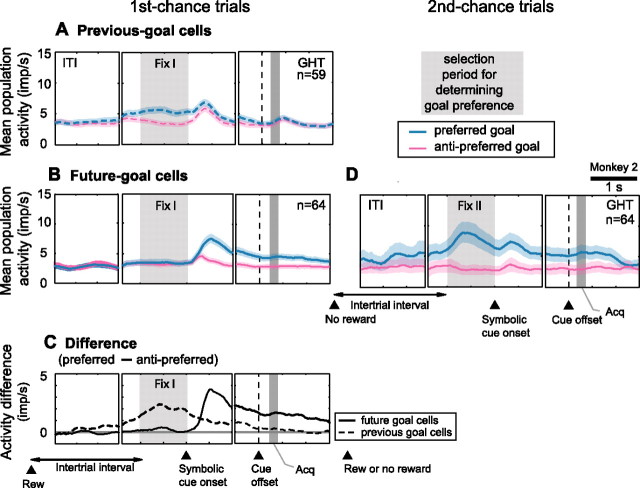
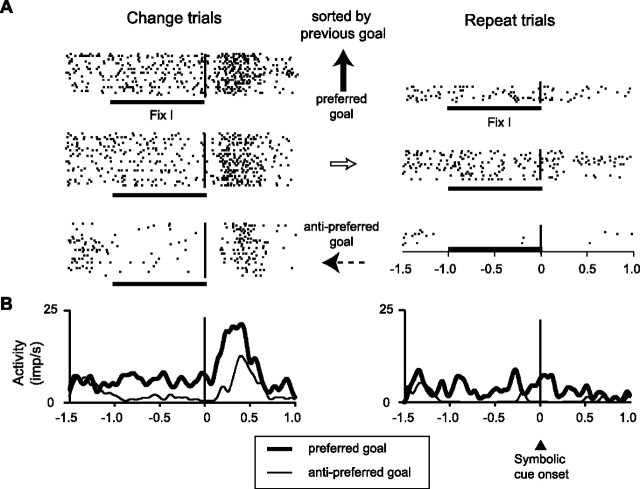
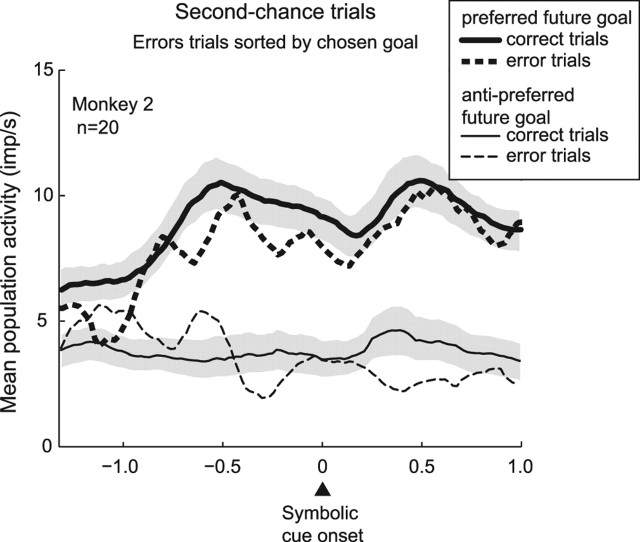
The primate prefrontal cortex plays a central role in choosing goals, along with a wide variety of additional functions, including short-term memory. In the present study, we examined neuronal activity in the prefrontal cortex as monkeys used abstract response strategies to select one of three spatial goals, a selection that depended on their memory of the most recent previous goal. During each trial, the monkeys selected a future goal on the basis of events from the previous trial, including both the symbolic visual cue that had appeared on that trial and the previous goal that the monkeys had selected. When a symbolic visual cue repeated from the previous trial, the monkeys stayed with their previous goal as the next (future) goal; when the cue changed, the monkeys shifted from their previous goal to one of the two remaining locations as their future goal. We found that prefrontal neurons had activity that reflected either previous goals or future goals, but only rarely did individual cells reflect both. This finding suggests that essentially separate neural networks encode these two aspects of spatial information processing. A failure to distinguish previous and future goals could lead to two kinds of maladaptive behavior. First, wrongly representing an accomplished goal as still pending could cause perseveration or compulsive checking, two disorders commonly attributed to dysfunction of the prefrontal cortex. Second, mistaking a pending goal as already accomplished could cause the failures of omission that occur commonly in dementia.
Figures
Similar articles
-
Neural Encoding of Self and Another Agent's Goal in the Primate Prefrontal Cortex: Human-Monkey Interactions.Cereb Cortex. 2016 Dec;26(12):4613-4622. doi: 10.1093/cercor/bhv224. Epub 2015 Oct 13. Cereb Cortex. 2016. PMID: 26464474
-
Delay-period activity in the primate prefrontal cortex encoding multiple spatial positions and their order of presentation.Behav Brain Res. 1997 Mar;84(1-2):203-23. doi: 10.1016/s0166-4328(96)00151-9. Behav Brain Res. 1997. PMID: 9079786
-
Neuronal activity in the primate prefrontal cortex in the process of motor selection based on two behavioral rules.J Neurophysiol. 2000 Apr;83(4):2355-73. doi: 10.1152/jn.2000.83.4.2355. J Neurophysiol. 2000. PMID: 10758139
-
The influence of recent decisions on future goal selection.Philos Trans R Soc Lond B Biol Sci. 2014 Nov 5;369(1655):20130477. doi: 10.1098/rstb.2013.0477. Philos Trans R Soc Lond B Biol Sci. 2014. PMID: 25267819 Free PMC article. Review.
-
Space representation in the prefrontal cortex.Prog Neurobiol. 2013 Apr;103:131-55. doi: 10.1016/j.pneurobio.2012.04.002. Epub 2012 Apr 11. Prog Neurobiol. 2013. PMID: 22521602 Review.
Cited by
-
What Happens When I Watch a Ballet and I Am Dyskinetic? A fMRI Case Report in Parkinson Disease.Front Psychol. 2020 Aug 7;11:1999. doi: 10.3389/fpsyg.2020.01999. eCollection 2020. Front Psychol. 2020. PMID: 32849162 Free PMC article.
-
Neural Circuits That Mediate Selective Attention: A Comparative Perspective.Trends Neurosci. 2018 Nov;41(11):789-805. doi: 10.1016/j.tins.2018.06.006. Epub 2018 Jul 31. Trends Neurosci. 2018. PMID: 30075867 Free PMC article. Review.
-
Attention-deficit/hyperactivity disorder (ADHD) in adults: a multilayered approach to a serious disorder of inattention to the future.Arq Neuropsiquiatr. 2024 Jul;82(7):1-12. doi: 10.1055/s-0044-1791513. Epub 2024 Oct 2. Arq Neuropsiquiatr. 2024. PMID: 39357854 Free PMC article. Review.
-
Compositional architecture: Orthogonal neural codes for task context and spatial memory in prefrontal cortex.bioRxiv [Preprint]. 2025 Feb 26:2025.02.25.640211. doi: 10.1101/2025.02.25.640211. bioRxiv. 2025. PMID: 40060470 Free PMC article. Preprint.
-
Static and dynamic coding in distinct cell types during associative learning in the prefrontal cortex.Nat Commun. 2023 Dec 14;14(1):8325. doi: 10.1038/s41467-023-43712-2. Nat Commun. 2023. PMID: 38097560 Free PMC article.
References
-
- Asaad WF, Rainer G, Miller EK (1998). Neural activity in the primate prefrontal cortex during associative learning. Neuron 21:1399–1407. - PubMed
-
- Asaad WF, Rainer G, Miller EK (2000). Task-specific neural activity in the primate prefrontal cortex. J Neurophysiol 84:451–459. - PubMed
-
- Barraclough DJ, Conroy ML, Lee D (2004). Prefrontal cortex and decision making in a mixed-strategy game. Nat Neurosci 7:404–410. - PubMed
-
- Brass M, Ruge H, Meiran N, Rubin O, Koch I, Zysset S, Prinz W, von Cramon DY (2003). When the same response has different meanings: recoding the response meaning in the lateral prefrontal cortex. NeuroImage 20:1026–1031. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources