

Secondary structure determines protein topology
- PMID: 16823044
- PMCID: PMC2242596
- DOI: 10.1110/ps.062305106
Secondary structure determines protein topology
Abstract
Using a test set of 13 small, compact proteins, we demonstrate that a remarkably simple protocol can capture native topology from secondary structure information alone, in the absence of long-range interactions. It has been a long-standing open question whether such information is sufficient to determine a protein's fold. Indeed, even the far simpler problem of reconstructing the three-dimensional structure of a protein from its exact backbone torsion angles has remained a difficult challenge owing to the small, but cumulative, deviations from ideality in backbone planarity, which, if ignored, cause large errors in structure. As a familiar example, a small change in an elbow angle causes a large displacement at the end of your arm; the longer the arm, the larger the displacement. Here, correct secondary structure assignments (alpha-helix, beta-strand, beta-turn, polyproline II, coil) were used to constrain polypeptide backbone chains devoid of side chains, and the most stable folded conformations were determined, using Monte Carlo simulation. Just three terms were used to assess stability: molecular compaction, steric exclusion, and hydrogen bonding. For nine of the 13 proteins, this protocol restricts the main chain to a surprisingly small number of energetically favorable topologies, with the native one prominent among them.
Figures
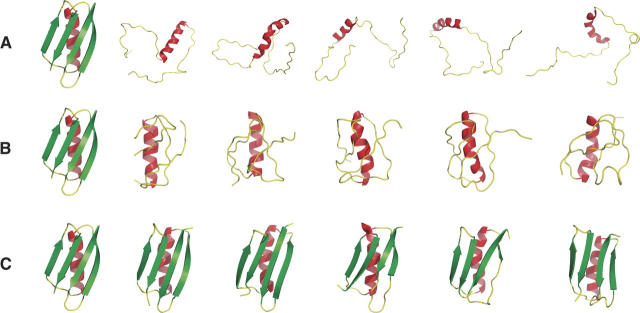

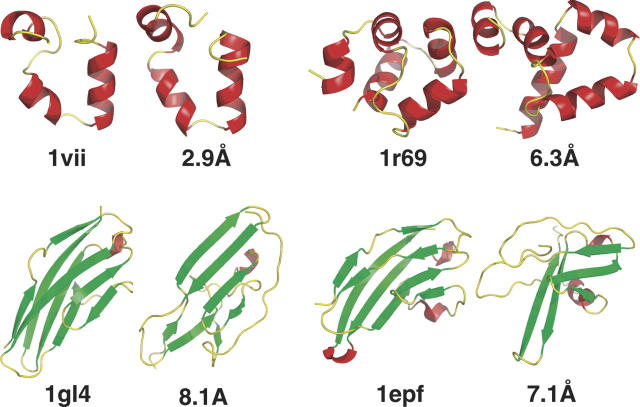
References
-
- Alexandrescu A.T. 2004. Strategy for supplementing structure calculations using limited data with hydrophobic distance restraints. Proteins 56: 117–129. - PubMed
-
- Anfinsen C.B. 1973. Principles that govern the folding of protein chains. Science 181: 223–230. - PubMed
-
- Baldwin R.L. and Rose G.D. 1999a. Is protein folding hierarchic? I: Local structure and peptide folding. Trends Biochem. Sci. 24: 26–33. - PubMed
-
- Baldwin R.L. and Rose G.D. 1999b. Is protein folding hierarchic? II: Folding intermediates and transition states. Trends Biochem. Sci. 24: 77–83. - PubMed
-
- Bradley P., Misura M.S., Baker D. 2005. Toward high-resolution de novo structure prediction for small proteins. Science 309: 1868–1871. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
