

The 5'-end transitional CpGs between the CpG islands and retroelements are hypomethylated in association with loss of heterozygosity in gastric cancers
- PMID: 16827945
- PMCID: PMC1552088
- DOI: 10.1186/1471-2407-6-180
The 5'-end transitional CpGs between the CpG islands and retroelements are hypomethylated in association with loss of heterozygosity in gastric cancers
Abstract
Background: A loss of heterozygosity (LOH) represents a unilateral chromosomal loss that reduces the dose of highly repetitive Alu, L1, and LTR retroelements. The aim of this study was to determine if the LOH events can affect the spread of retroelement methylation in the 5'-end transitional area between the CpG islands and their nearest retroelements.
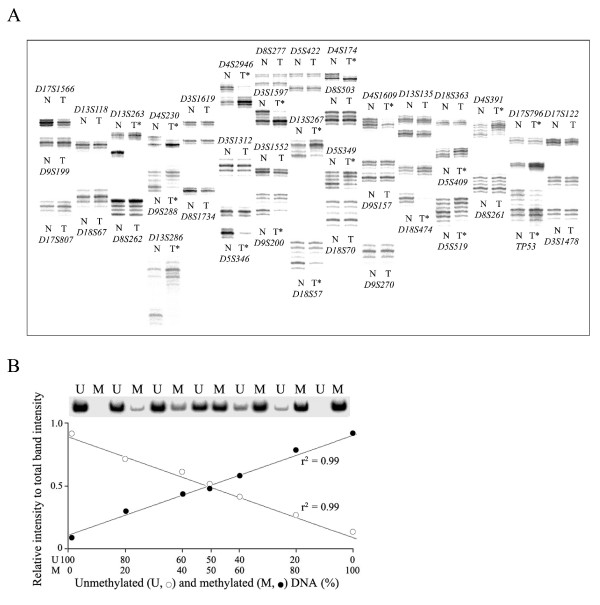
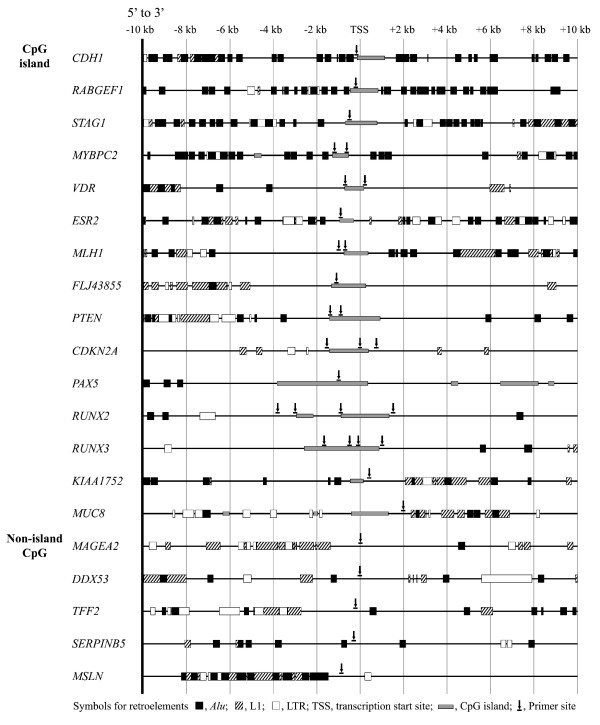
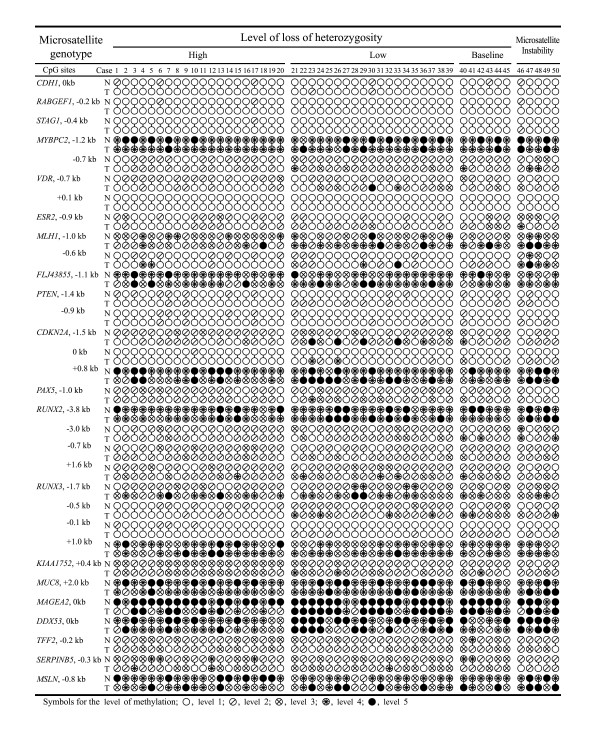
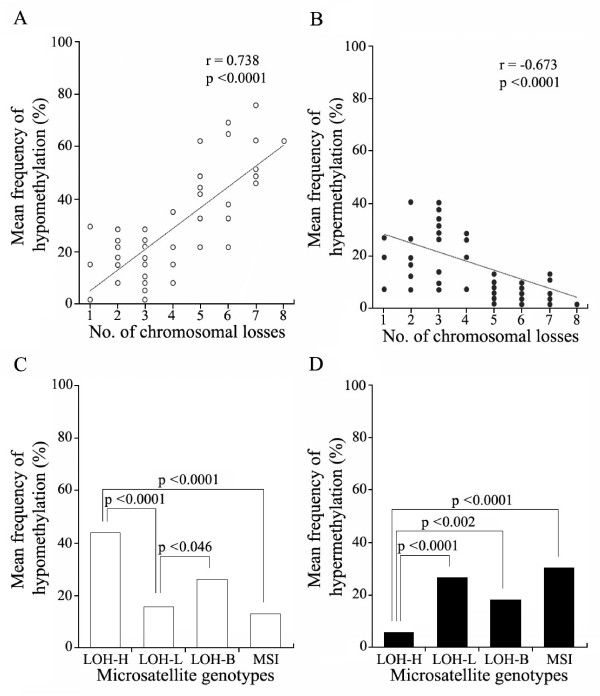
Methods: The 5'-transitional area of all human genes (22,297) was measured according to the nearest retroelements to the transcription start sites. For 50 gastric cancer specimens, the level of LOH events on eight cancer-associated chromosomes was estimated using the microsatellite markers, and the 5'-transitional CpGs of 20 selected genes were examined by methylation analysis using the bisulfite-modified DNA.
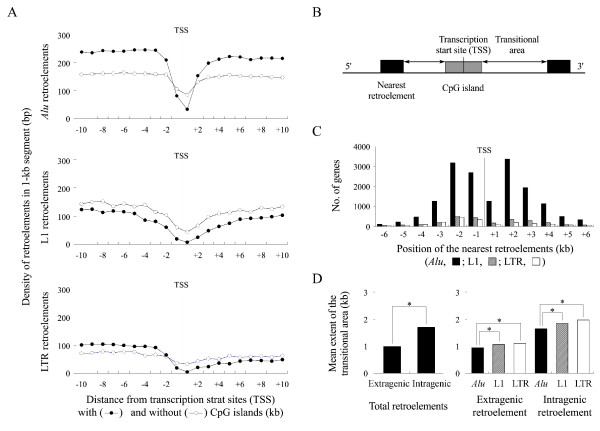
Results: The extent of the transitional area was significantly shorter with the nearest Alu elements than with the nearest L1 and LTR elements, as well as in the extragenic regions containing a higher density of retroelements than in the intragenic regions. The CpG islands neighbouring a high density of Alu elements were consistently hypomethylated in both normal and tumor tissues. The 5'-transitional methylated CpG sites bordered by a low density of Alu elements or the L1 and LTR elements were hypomethylated more frequently in the high-level LOH cases than in the low-level LOH cases.
Conclusion: The 5'-transitional methylated CpG sites not completely protected by the Alu elements were hypomethylated in association with LOH events in gastric cancers. This suggests that an irreversible unbalanced decrease in the genomic dose reduces the spread of L1 methylation in the 5'-end regions of genes.
Figures

References
-
- Vogelstein B, Fearon ER, Kern SE, Hamilton SR, Preisinger AC, Nakamura Y, White R. Allelotype of colorectal carcinomas. Science. 1989;244:207–211. - PubMed
-
- Choi SW, Lee KJ, Bae YA, Min KO, Kwon MS, Kim KM, Rhyu MG. Genetic classification of colorectal cancer based on chromosomal loss and microsatellite instability predicts survival. Clin Cancer Res. 2002;8:2311–2322. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
