

Review
doi: 10.1038/nrm1961.
The biogenesis and regulation of telomerase holoenzymes
Affiliations
- PMID: 16829980
- PMCID: PMC2915765
- DOI: 10.1038/nrm1961
Item in Clipboard
Review
The biogenesis and regulation of telomerase holoenzymes
Nat Rev Mol Cell Biol.
2006 Jul.
Abstract
Chromosome stability requires a dynamic balance of DNA loss and gain in each terminal tract of telomeric repeats. Repeat addition by a specialized reverse transcriptase, telomerase, has an important role in maintaining this equilibrium. Insights that have been gained into the cellular pathways for biogenesis and regulation of telomerase ribonucleoproteins raise new questions, particularly concerning the dynamic nature of this unique polymerase.
Figures
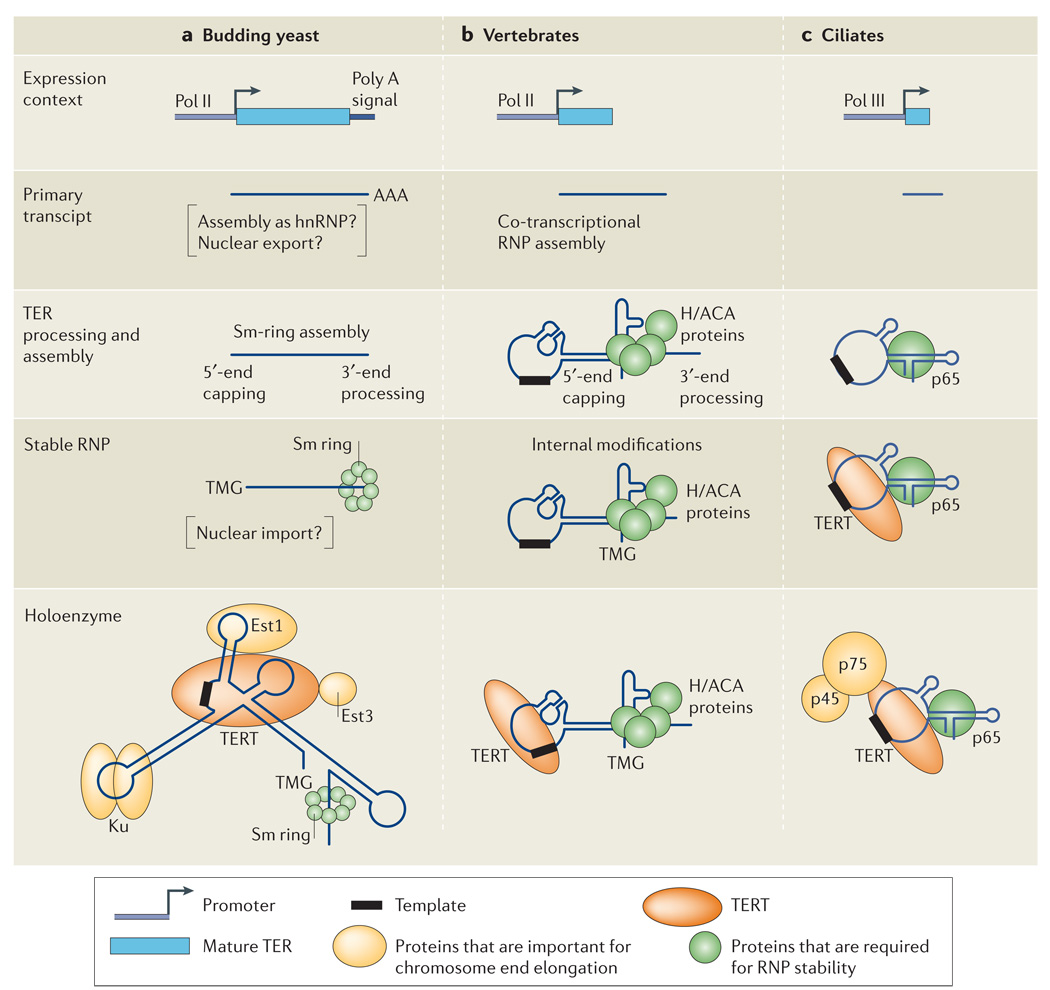
Schematic illustration of the events that accompany telomerase biogenesis in vivo through the following steps: telomerase RNA subunit (TER) expression, TER processing and assembly to form a stable telomerase ribonucleoprotein (RNP), and additional assembly to form a telomerase holoenzyme(s). a | In budding yeast, the TER precursor is transcribed by RNA polymerase II (Pol II) and gains a tract of polyadenosine (Poly A). It then undergoes assembly steps (possibly including heterogeneous nuclear (hn) RNP assembly and nuclear export) that result in the loading of a heteroheptameric ring of Sm proteins and the addition of the 5′-trimethylguanosine (TMG) cap. Holoenzyme form(s) of RNP also include the telomerase subunits Est1 and Est3 as well as telomerase reverse transcriptase (TERT) and the DNA-end-binding protein Ku. b | In human cells (vertebrates in the figure), the TER precursor is transcribed by Pol II. The nascent transcript is modified with a 5′-TMG cap and binds to the H/ACA-motif proteins that direct 3′-end processing. Holoenzyme form(s) of RNP also include TERT. c | In Tetrahymena thermophila (ciliates in the figure), TER is transcribed by Pol III and binds to the telomerase subunits p65 and TERT. Holoenzyme form(s) of RNP also include telomerase p45 and p75.
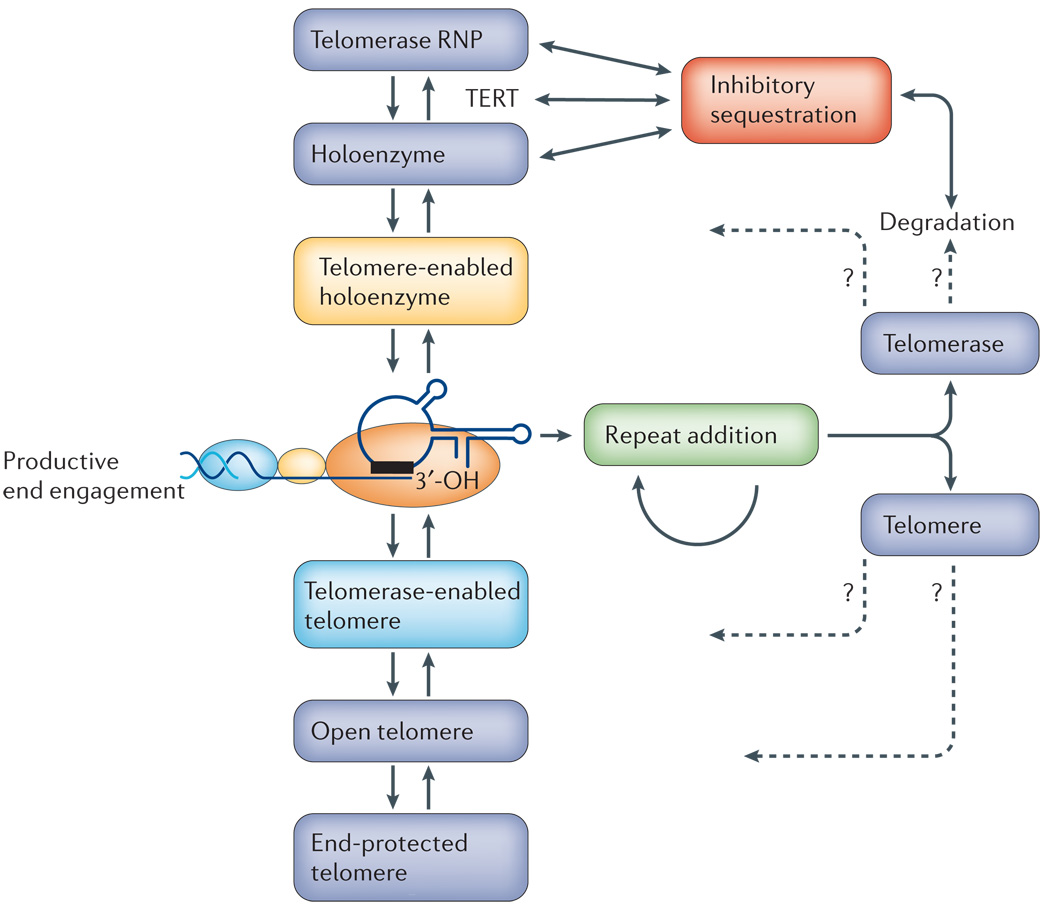
Telomerase engages with telomeres in a dynamic manner that is governed by changes in the state of both interaction partners. The dark blue strand of DNA (middle left) represents guanosine-rich telomeric repeats and has the chromosome 3′-OH end, which must be positioned in the telomerase-holoenzyme active site for elongation. To reach this state, telomerase ribonucleoprotein (RNP) assembles with the telomerase reverse transcriptase (TERT), to generate a catalytically active enzyme, and with additional factors to ultimately form an enzyme that has activity on telomeres. This process seems to be affected by the sequestration of components in distinct subnuclear compartments. To be elongated by telomerase, telomeric chromatin is remodelled into a more accessible state and binds to factors that promote telomerase interaction. After each telomeric repeat synthesis, telomerase either repositions itself for processive repeat synthesis or it dissociates from the telomere. Dissociation might involve reversible disassembly or it might require the remodelling or degradation of the telomerase holoenzyme and/or the telomeric chromatin.
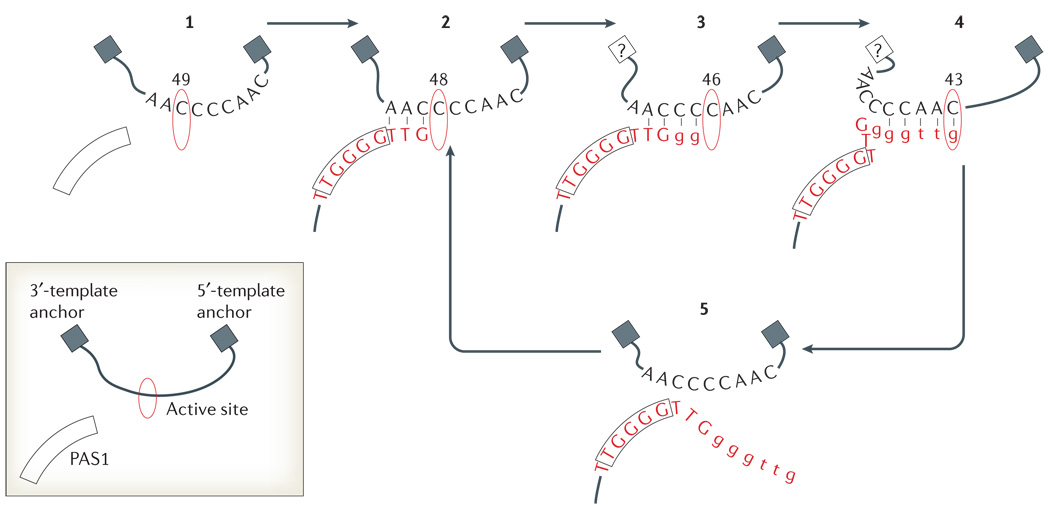
The telomerase catalytic cycle is illustrated based on studies of the Tetrahymena thermophila holoenzyme. The telomerase RNA subunit (TER) template is shown with limited 5′- and 3′-flanking regions that anchor it in the vicinity of the active site; the template-adjacent single-stranded-DNA-binding site PAS1 (primer/product alignment/anchor site-1) is also shown (inset box). 1 | Template positioning at default is thought to place residue 49 of TER in the active site. 2 | Primer binding at PAS1 and 3′-end hybridization with the template can reposition the template from this default position. 3 | Elongation is proposed to occur such that up to five base-pairs of hybrid can form between the template and the product. 4 | Some hybrid dissociation occurs before the 5′ end of the template enters the active site, (5) facilitating product release from the template. The free template can relax into the default position and (2) can re-engage a product 3′ end for processive repeat synthesis.
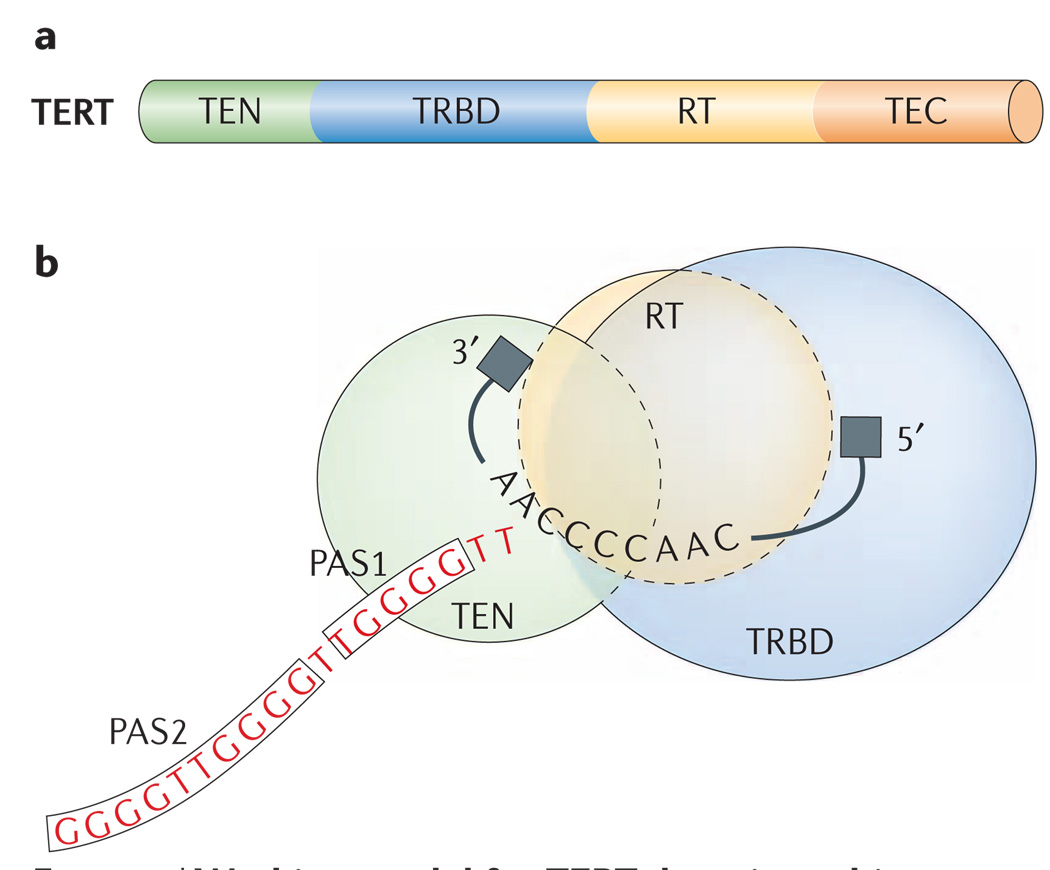
a | The telomerase reverse transcriptase (TERT) is divided into four regions, the TERT essential N-terminal (TEN) domain, the TERT high-affinity RNA-binding domain (TRBD), the reverse-transcriptase (RT) motifs and the TERT C-terminal (TEC) extension. b | A speculative illustration of the three TERT domains, TEN, TRBD and RT, that shows the coordination of the proposed sites of protein–nucleic-acid interaction in Tetrahymena thermophila telomerase. The template is shown with limited 5′- and 3′-flanking regions, as in FIG. 3. Single-stranded DNA that is adjacent to the template binds to PAS1 (primer/product alignment/anchor site-1), with additional PAS2 contacts in a subset of holoenzymes. At least part of PAS1 is located in the TEN domain, at least part of the 5′-template anchor site is located in the TRBD domain, and the active site is located in the RT domain. A 3′-template anchor site is modelled as located in the TEN domain, but the principles of 3′-template anchoring are not well understood. The TEN and TEC domains could both be part of the PAS2 (not shown; see text for discussion).
References
-
- Blackburn EH, Gall JG. A tandemly repeated sequence at the termini of the extrachromosomal ribosomal RNA genes in Tetrahymena. J. Mol. Biol. 1978;120:33–53. - PubMed
-
- de Lange T. Shelterin: the protein complex that shapes and safeguards human telomeres. Genes Dev. 2005;19:2100–2110. - PubMed
-
- Bhattacharyya MK, Lustig AJ. Telomere dynamics in genome stability. Trends Biochem. Sci. 2006;31:114–122. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
