

Pre-Bötzinger complex: a brainstem region that may generate respiratory rhythm in mammals
- PMID: 1683005
- PMCID: PMC3209964
- DOI: 10.1126/science.1683005
Pre-Bötzinger complex: a brainstem region that may generate respiratory rhythm in mammals
Abstract
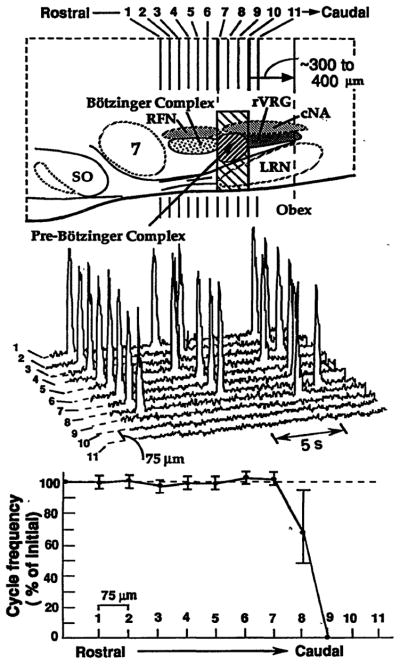
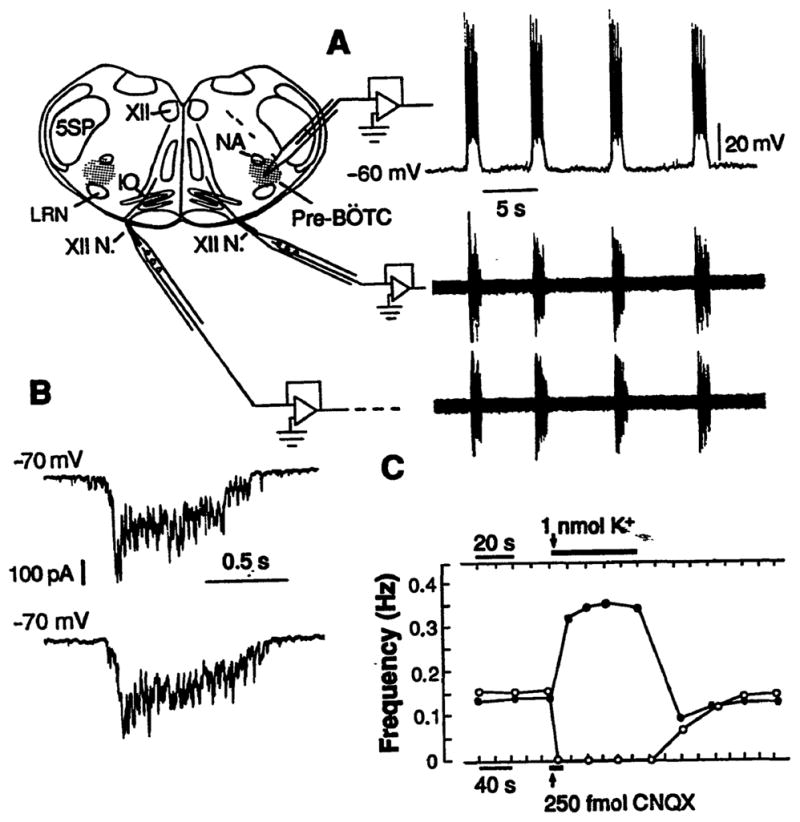
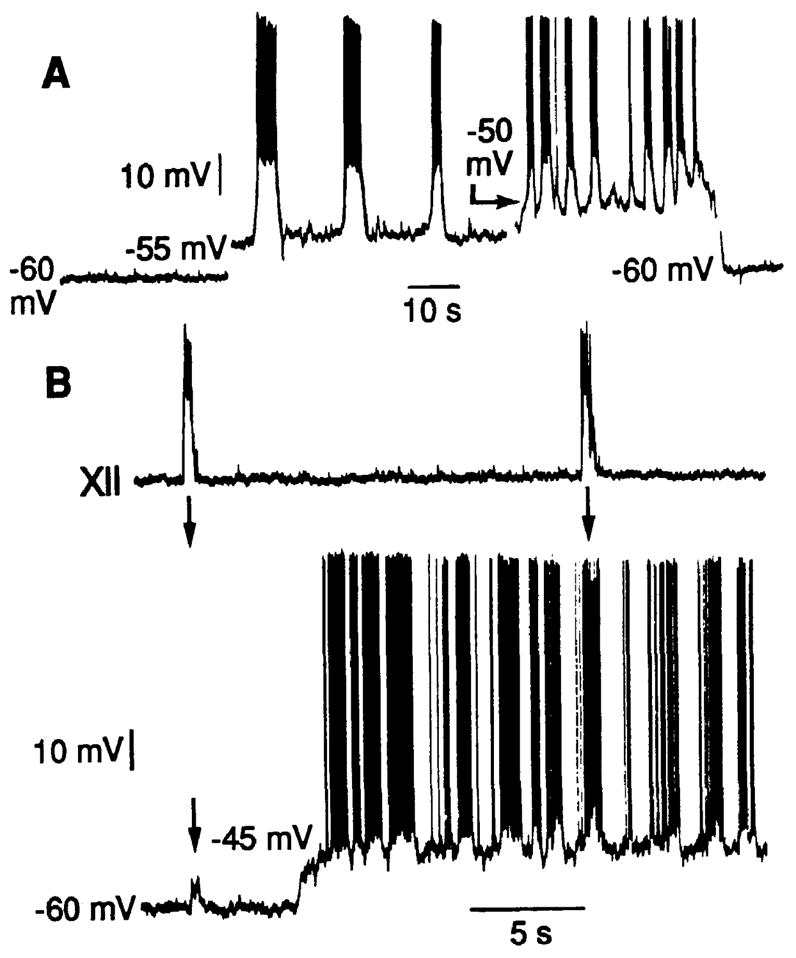
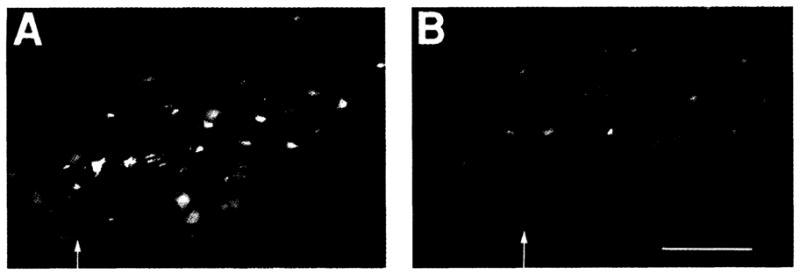
The location of neurons generating the rhythm of breathing in mammals is unknown. By microsection of the neonatal rat brainstem in vitro, a limited region of the ventral medulla (the pre-Bötzinger Complex) that contains neurons essential for rhythmogenesis was identified. Rhythm generation was eliminated by removal of only this region. Medullary slices containing the pre-Bötzinger Complex generated respiratory-related oscillations similar to those generated by the whole brainstem in vitro, and neurons with voltage-dependent pacemaker-like properties were identified in this region. Thus, the respiratory rhythm in the mammalian neonatal nervous system may result from a population of conditional bursting pacemaker neurons in the pre-Bötzinger Complex.
Figures
References
-
- Fluorens P. C R Acad Sci (Paris) 1851;33:437.
-
- Smith JC, Greer JJ, Liu G, Feldman JL. J Neurophysiol. 1990;64:1149. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
