

Pheromone detection in male mice depends on signaling through the type 3 adenylyl cyclase in the main olfactory epithelium
- PMID: 16837584
- PMCID: PMC6674185
- DOI: 10.1523/JNEUROSCI.1967-06.2006
Pheromone detection in male mice depends on signaling through the type 3 adenylyl cyclase in the main olfactory epithelium
Abstract
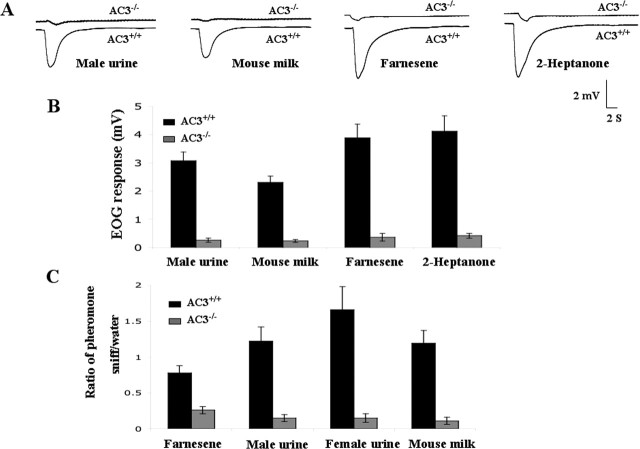
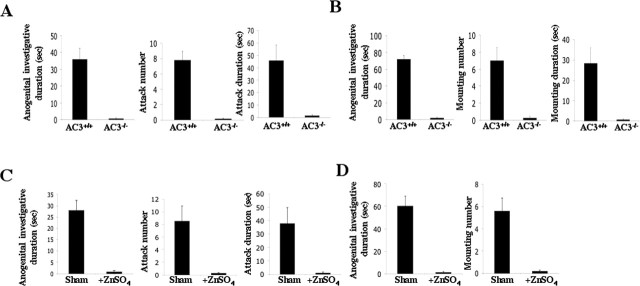
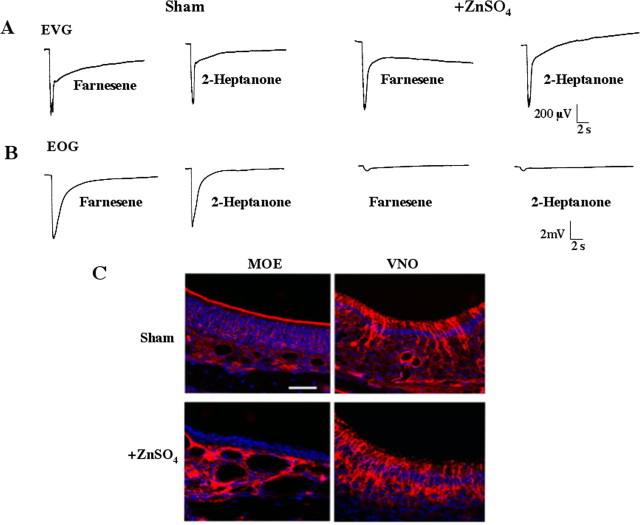
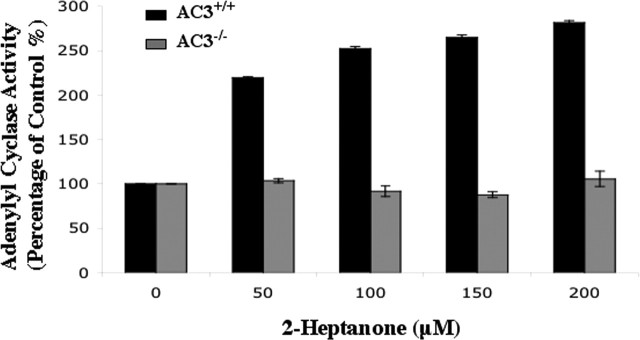
Terrestrial vertebrates have evolved two anatomically and mechanistically distinct chemosensory structures: the main olfactory epithelium (MOE) and the vomeronasal organ (VNO). Although it has been generally thought that pheromones are detected through the VNO, whereas other chemicals are sensed by the MOE, recent evidence suggests that some pheromones may be detected through the MOE. Odorant receptors in the MOE are coupled to the type 3 adenylyl cyclase (AC3), an enzyme not expressed in the VNO. Consequently, odorants and pheromones do not elicit electrophysiological responses in the MOE of AC3-/- mice, although VNO function is intact. Here we report that AC3-/- mice cannot detect mouse milk, urine, or mouse pheromones. Inter-male aggressiveness and male sexual behaviors are absent in AC3-/- mice. Furthermore, adenylyl cyclase activity in membranes prepared from the MOE of wild-type mice, but not AC3-/- mice, is stimulated by 2-heptanone, a mouse pheromone. We conclude that signaling through AC3 in the MOE is obligatory for male sexual behavior, male-male aggressiveness, and the detection of some pheromones.
Figures
Similar articles
-
Vomeronasal organ detects odorants in absence of signaling through main olfactory epithelium.Nat Neurosci. 2003 May;6(5):519-25. doi: 10.1038/nn1039. Nat Neurosci. 2003. PMID: 12665798
-
Detection of odorants through the main olfactory epithelium and vomeronasal organ of mice.Nutr Rev. 2004 Nov;62(11 Pt 2):S189-92; discussion S224-41. doi: 10.1111/j.1753-4887.2004.tb00098.x. Nutr Rev. 2004. PMID: 15630934 Review.
-
Are pheromones detected through the main olfactory epithelium?Mol Neurobiol. 2007 Jun;35(3):317-23. doi: 10.1007/s12035-007-0014-1. Mol Neurobiol. 2007. PMID: 17917120 Review.
-
Maternal behavior is impaired in female mice lacking type 3 adenylyl cyclase.Neuropsychopharmacology. 2011 Mar;36(4):772-81. doi: 10.1038/npp.2010.211. Epub 2010 Dec 8. Neuropsychopharmacology. 2011. PMID: 21150908 Free PMC article.
-
Sensory transduction in vomeronasal neurons: evidence for G alpha o, G alpha i2, and adenylyl cyclase II as major components of a pheromone signaling cascade.J Neurosci. 1996 Feb 1;16(3):909-18. doi: 10.1523/JNEUROSCI.16-03-00909.1996. J Neurosci. 1996. PMID: 8558259 Free PMC article.
Cited by
-
Neural circuits of social behaviors: Innate yet flexible.Neuron. 2021 May 19;109(10):1600-1620. doi: 10.1016/j.neuron.2021.02.012. Epub 2021 Mar 10. Neuron. 2021. PMID: 33705708 Free PMC article. Review.
-
Deletion of Type 3 Adenylyl Cyclase Perturbs the Postnatal Maturation of Olfactory Sensory Neurons and Olfactory Cilium Ultrastructure in Mice.Front Cell Neurosci. 2017 Jan 19;11:1. doi: 10.3389/fncel.2017.00001. eCollection 2017. Front Cell Neurosci. 2017. PMID: 28154525 Free PMC article.
-
Regulation of adult neurogenesis by behavior and age in the accessory olfactory bulb.Mol Cell Neurosci. 2011 Aug;47(4):274-85. doi: 10.1016/j.mcn.2011.05.003. Epub 2011 May 10. Mol Cell Neurosci. 2011. PMID: 21600286 Free PMC article.
-
Stimulation of electro-olfactogram responses in the main olfactory epithelia by airflow depends on the type 3 adenylyl cyclase.J Neurosci. 2012 Nov 7;32(45):15769-78. doi: 10.1523/JNEUROSCI.2180-12.2012. J Neurosci. 2012. PMID: 23136416 Free PMC article.
-
Autistic-like behaviour in Scn1a+/- mice and rescue by enhanced GABA-mediated neurotransmission.Nature. 2012 Sep 20;489(7416):385-90. doi: 10.1038/nature11356. Epub 2012 Aug 22. Nature. 2012. PMID: 22914087 Free PMC article.
References
-
- Bahrke MS, Yesalis CE III, Wright JE (1996). Psychological and behavioural effects of endogenous testosterone and anabolic-androgenic steroids. An update. Sports Med 22:367–390. - PubMed
-
- Belluscio L, Gold GH, Nemes A, Axel R (1998). Mice deficient in G(olf) are anosmic. Neuron 20:69–81. - PubMed
-
- Brain PF, Haug M (1992). Hormonal and neurochemical correlates of various forms of animal “aggression.”. Psychoneuroendocrinology 17:537–551. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases