

Effects of potent inhibitors of the retinoid cycle on visual function and photoreceptor protection from light damage in mice
- PMID: 16837623
- PMCID: PMC4106413
- DOI: 10.1124/mol.106.026823
Effects of potent inhibitors of the retinoid cycle on visual function and photoreceptor protection from light damage in mice
Abstract
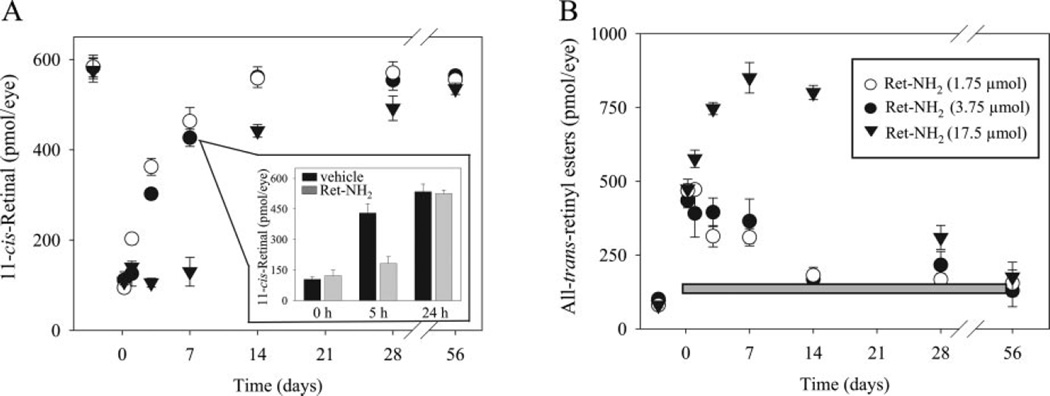
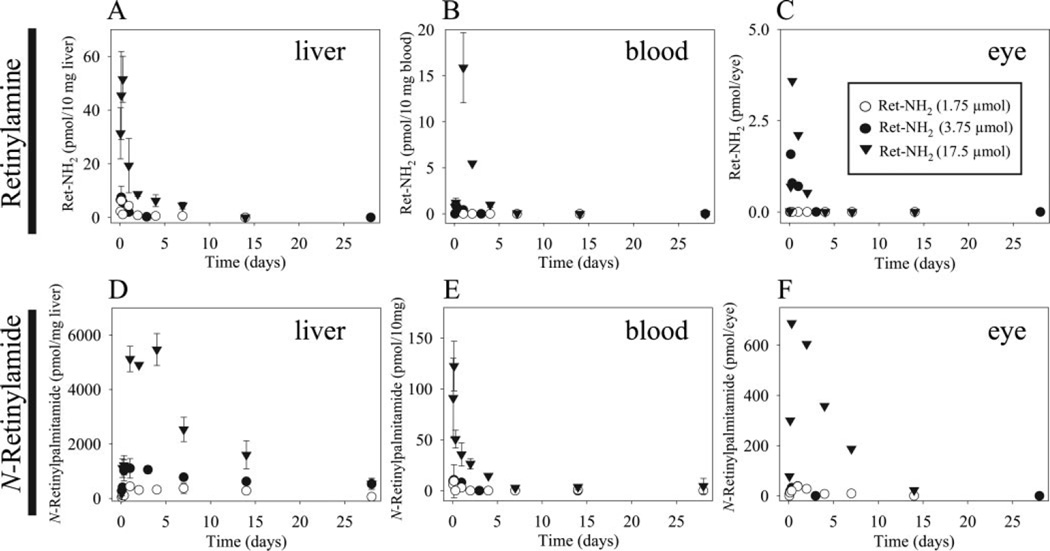
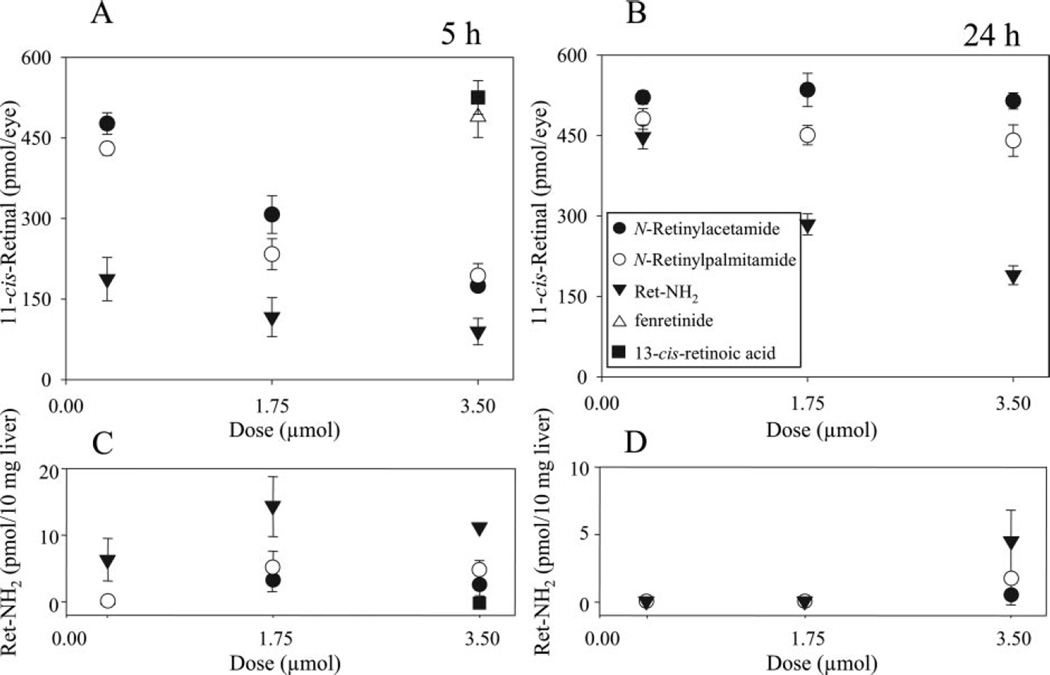
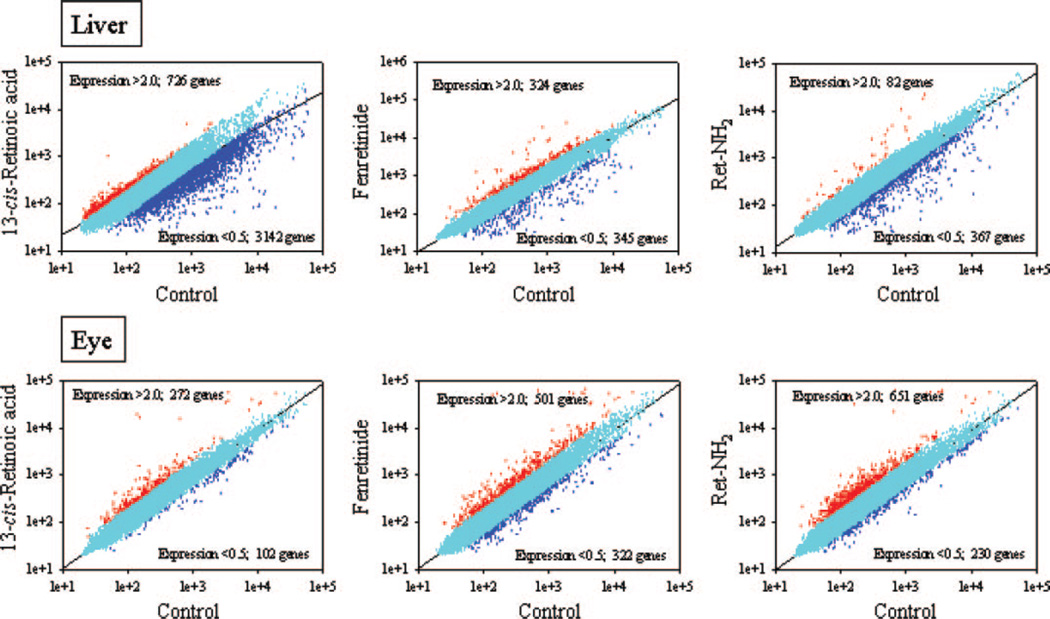
Regeneration of the chromophore 11-cis-retinal is essential for the generation of light-sensitive visual pigments in the vertebrate retina. A deficiency in 11-cis-retinal production leads to congenital blindness in humans; however, a buildup of the photoisomerized chromophore can also be detrimental. Such is the case when the photoisomerized all-trans-retinal is produced but cannot be efficiently cleared from the internal membrane of the outer segment discs. Sustained increase of all-trans-retinal can lead to the formation of toxic condensation products in the eye. Thus, there is a need for potent, selective inhibitors that can regulate the flux of retinoids through the metabolism pathway termed the visual (retinoid) cycle. Here we systematically study the effects of the most potent inhibitor of this cycle, retinylamine (Ret-NH2), on visual function in mice. Prolonged, sustainable, but reversible suppression of the visual function was observed by Ret-NH2 as a result of its storage in a prodrug form, N-retinylamides. Direct comparison of other inhibitors such as fenretinide and 13-cis-retinoic acid showed multiple advantages of Ret-NH2 and its amides, including a higher potency, specificity, and lower transcription activation. Our results also revealed that mice treated with Ret-NH2 were completely resistant to the light-induced retina damage. As an experimental tool, Ret-NH2 allows the replacement of the native chromophore with synthetic analogs in wild-type mice to better understand the function of the chromophore in the activation of rhodopsin and its metabolism through the retinoid cycle.
Figures
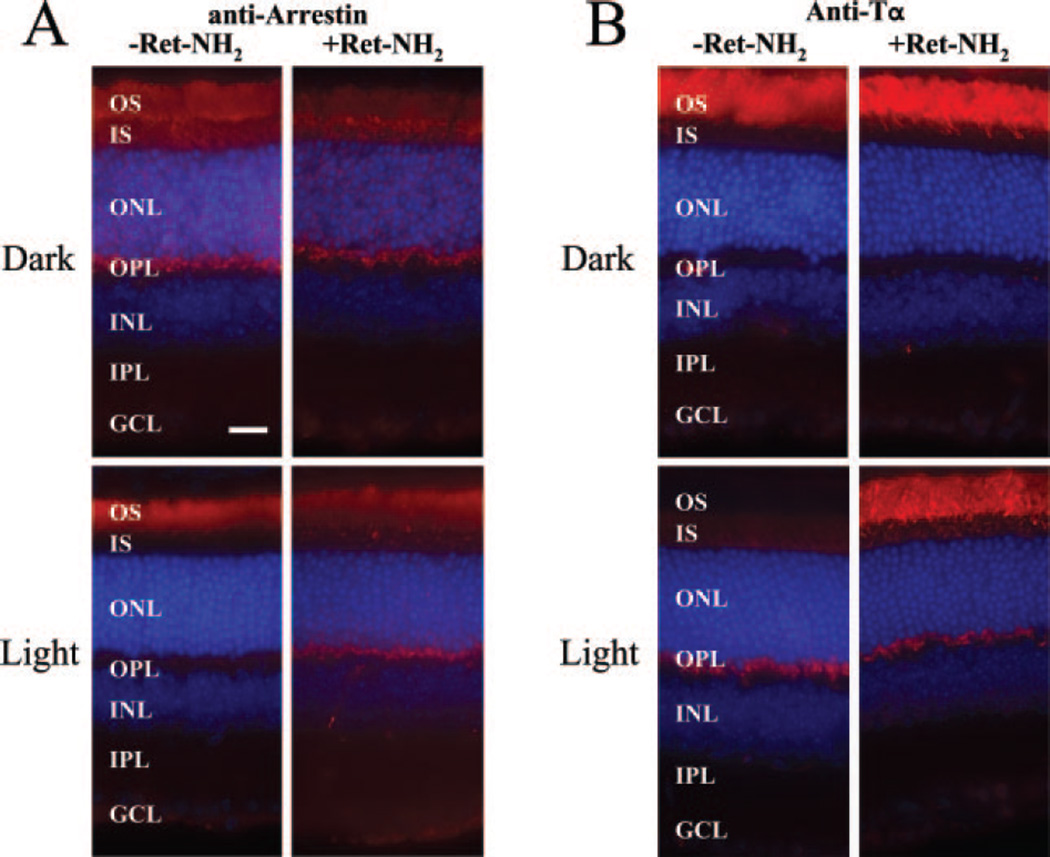
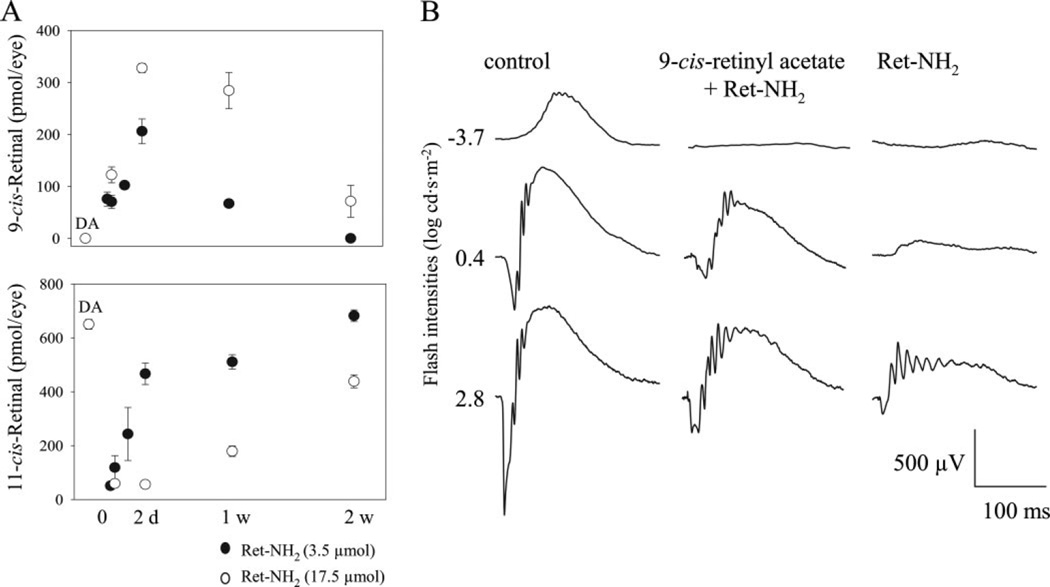
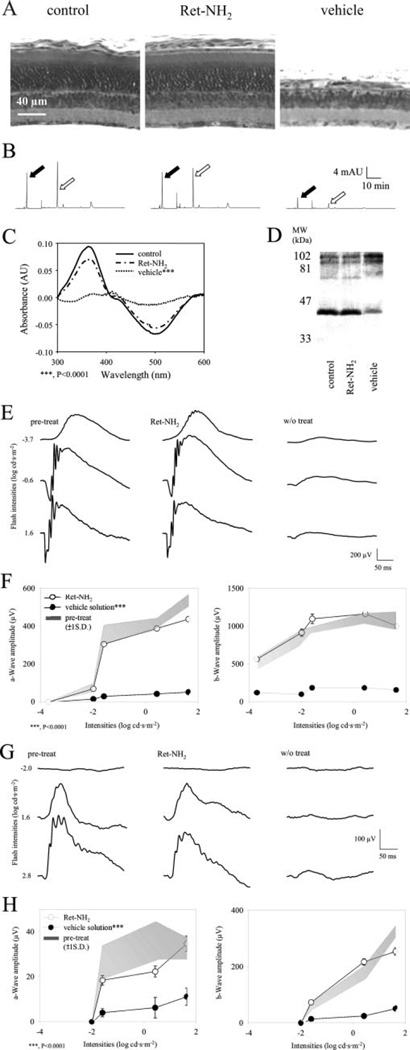
Similar articles
-
Positively charged retinoids are potent and selective inhibitors of the trans-cis isomerization in the retinoid (visual) cycle.Proc Natl Acad Sci U S A. 2005 Jun 7;102(23):8162-7. doi: 10.1073/pnas.0503318102. Epub 2005 May 25. Proc Natl Acad Sci U S A. 2005. PMID: 15917330 Free PMC article.
-
Lecithin:retinol acyltransferase is responsible for amidation of retinylamine, a potent inhibitor of the retinoid cycle.J Biol Chem. 2005 Dec 23;280(51):42263-73. doi: 10.1074/jbc.M509351200. Epub 2005 Oct 10. J Biol Chem. 2005. PMID: 16216874
-
Metabolic basis of visual cycle inhibition by retinoid and nonretinoid compounds in the vertebrate retina.J Biol Chem. 2008 Apr 11;283(15):9543-54. doi: 10.1074/jbc.M708982200. Epub 2008 Jan 14. J Biol Chem. 2008. PMID: 18195010 Free PMC article.
-
Key enzymes of the retinoid (visual) cycle in vertebrate retina.Biochim Biophys Acta. 2012 Jan;1821(1):137-51. doi: 10.1016/j.bbalip.2011.03.005. Epub 2011 Apr 5. Biochim Biophys Acta. 2012. PMID: 21447403 Free PMC article. Review.
-
Confronting complexity: the interlink of phototransduction and retinoid metabolism in the vertebrate retina.Prog Retin Eye Res. 2001 Jul;20(4):469-529. doi: 10.1016/s1350-9462(01)00002-7. Prog Retin Eye Res. 2001. PMID: 11390257 Review.
Cited by
-
Visual Cycle Modulation as an Approach toward Preservation of Retinal Integrity.PLoS One. 2015 May 13;10(5):e0124940. doi: 10.1371/journal.pone.0124940. eCollection 2015. PLoS One. 2015. PMID: 25970164 Free PMC article.
-
Noninvasive two-photon microscopy imaging of mouse retina and retinal pigment epithelium through the pupil of the eye.Nat Med. 2014 Jul;20(7):785-9. doi: 10.1038/nm.3590. Epub 2014 Jun 22. Nat Med. 2014. PMID: 24952647 Free PMC article.
-
PBN (Phenyl-N-Tert-Butylnitrone)-Derivatives Are Effective in Slowing the Visual Cycle and Rhodopsin Regeneration and in Protecting the Retina from Light-Induced Damage.PLoS One. 2015 Dec 22;10(12):e0145305. doi: 10.1371/journal.pone.0145305. eCollection 2015. PLoS One. 2015. PMID: 26694648 Free PMC article.
-
Advances and therapeutic opportunities in visual cycle modulation.Prog Retin Eye Res. 2025 May;106:101360. doi: 10.1016/j.preteyeres.2025.101360. Epub 2025 Apr 23. Prog Retin Eye Res. 2025. PMID: 40280538 Free PMC article. Review.
-
Abnormal Cannabidiol Modulates Vitamin A Metabolism by Acting as a Competitive Inhibitor of CRBP1.ACS Chem Biol. 2019 Mar 15;14(3):434-448. doi: 10.1021/acschembio.8b01070. Epub 2019 Feb 18. ACS Chem Biol. 2019. PMID: 30721022 Free PMC article.
References
-
- Allikmets R, Singh N, Sun H, Shroyer NF, Hutchinson A, Chidambaram A, Gerrard B, Baird L, Stauffer D, Peiffer A, et al. A photoreceptor cell-specific ATP-binding transporter gene (ABCR) is mutated in recessive Stargardt macular dystrophy. Nat Genet. 1997;15:236–246. - PubMed
-
- Beharry S, Zhong M, Molday RS. N-retinylidene-phosphatidylethanolamine is the preferred retinoid substrate for the photoreceptor-specific ABC transporter ABCA4 (ABCR) J Biol Chem. 2004;279:53972–53979. - PubMed
-
- Chambon P. A decade of molecular biology of retinoic acid receptors. FASEB J. 1996;10:940–954. - PubMed
-
- Golczak M, Imanishi Y, Kuksa V, Maeda T, Kubota R, Palczewski K. Lecithin:retinol acyltransferase is responsible for amidation of retinylamine, a potent inhibitor of the retinoid cycle. J Biol Chem. 2005a;280:42263–42273. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
