

Drosophila CENP-A mutations cause a BubR1-dependent early mitotic delay without normal localization of kinetochore components
- PMID: 16839185
- PMCID: PMC1500813
- DOI: 10.1371/journal.pgen.0020110
Drosophila CENP-A mutations cause a BubR1-dependent early mitotic delay without normal localization of kinetochore components
Abstract
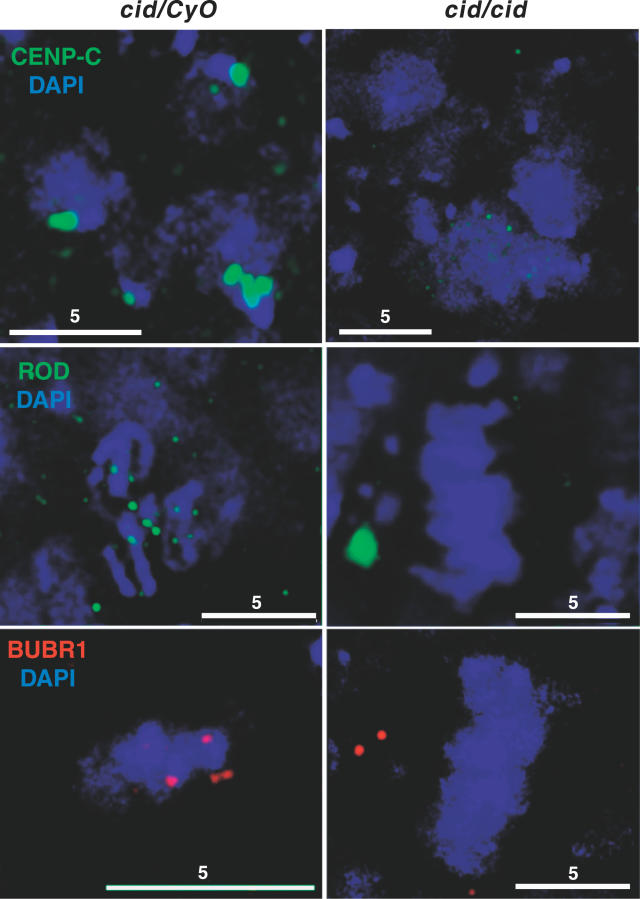
The centromere/kinetochore complex plays an essential role in cell and organismal viability by ensuring chromosome movements during mitosis and meiosis. The kinetochore also mediates the spindle attachment checkpoint (SAC), which delays anaphase initiation until all chromosomes have achieved bipolar attachment of kinetochores to the mitotic spindle. CENP-A proteins are centromere-specific chromatin components that provide both a structural and a functional foundation for kinetochore formation. Here we show that cells in Drosophila embryos homozygous for null mutations in CENP-A (CID) display an early mitotic delay. This mitotic delay is not suppressed by inactivation of the DNA damage checkpoint and is unlikely to be the result of DNA damage. Surprisingly, mutation of the SAC component BUBR1 partially suppresses this mitotic delay. Furthermore, cid mutants retain an intact SAC response to spindle disruption despite the inability of many kinetochore proteins, including SAC components, to target to kinetochores. We propose that SAC components are able to monitor spindle assembly and inhibit cell cycle progression in the absence of sustained kinetochore localization.
Conflict of interest statement
Competing interests. The authors have declared that no competing interests exist.
Figures






References
-
- Cleveland DW, Mao Y, Sullivan KF. Centromeres and kinetochores: From epigenetics to mitotic checkpoint signaling. Cell. 2003;112:407–421. - PubMed
-
- Rieder CL, Maiato H. Stuck in division or passing through: What happens when cells cannot satisfy the spindle assembly checkpoint. Dev Cell. 2004;7:637–651. - PubMed
-
- Michel LS, Liberal V, Chatterjee A, Kirchwegger R, Pasche B, et al. MAD2 haplo-insufficiency causes premature anaphase and chromosome instability in mammalian cells. Nature. 2001;409:355–359. - PubMed
-
- Grady WM. Genomic instability and colon cancer. Cancer Metastasis Rev. 2004;23:11–27. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
