

doi: 10.1128/JVI.00435-06.
Surface mutagenesis of the bovine papillomavirus E1 DNA binding domain reveals residues required for multiple functions related to DNA replication
Affiliations
- PMID: 16840329
- PMCID: PMC1563737
- DOI: 10.1128/JVI.00435-06
Item in Clipboard
Surface mutagenesis of the bovine papillomavirus E1 DNA binding domain reveals residues required for multiple functions related to DNA replication
J Virol.
2006 Aug.
Abstract
The E1 protein from papillomaviruses is a multifunctional protein with complex functions required for the initiation of viral DNA replication. We have performed a surface mutagenesis of the well-characterized E1 DNA binding domain (DBD). We demonstrate that substitutions of multiple residues on the surface of the E1 DBD are defective for DNA replication without affecting the DNA binding activity of the protein. The defects of individual substitutions include failure to form the double trimer that melts the ori and failure to form the double hexamer that unwinds the ori. These results demonstrate that the DBD plays an essential role in multiple DNA replication-related processes apart from DNA binding.
Figures
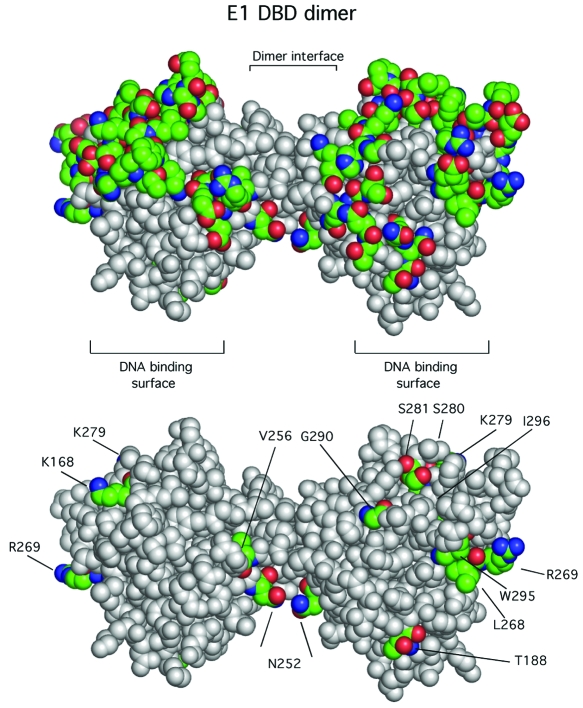
Space-filling representation of the E1 DBD dimer as determined by X-ray crystallography. The colored residues in panel A represent the 63 residues on the surface of the E1 DBD that were changed to alanines. In panel B, the 12 residues that had a significant deleterious effect on DNA replication when changed to alanine are shown.
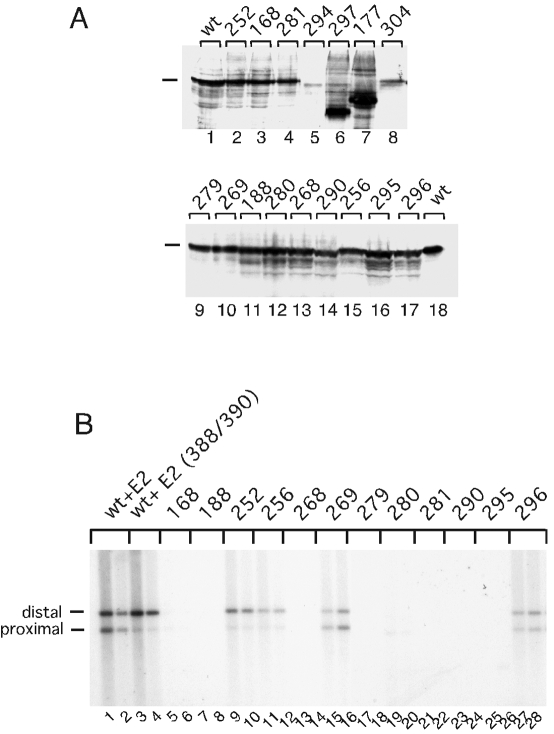
(A) In vivo expression of mutant E1 proteins. The E1 expression vectors that were unable to support DNA replication in vivo were transfected into COS cells. Forty-eight hours after transfection, the cells were lysed by the addition of Laemmli-loading dye and the resulting lysates were analyzed by SDS-PAGE, transferred to nitrocellulose, and probed with a monoclonal E1 antibody. (B) In vivo replication assays. Replication assays were carried out by transfection of the E1 and E2 expression vectors pCGE1 and pCGE2, together with the ori plasmids X/12/X (proximal E2 BS) and 11/X/X (distal E2 BS) (25). In lanes 1 and 2, wt E1 and wt E2 were used. In lanes 3 and 4, wt E1 and the mutant E2 (388/390) (13) were used. In lanes 5 through 28, the respective E1 mutants were cotransfected with wt E2. Low-molecular-weight DNA was harvested 48 and 60 h after transfection, digested with DpnI, EcoO109I, and HindIII, run on agarose gels, transferred to nitrocellulose, and probed with an ori probe. After digestion with EcoO109I and HindIII, the ori plasmid with the proximal E2 BS gives rise to a 2.5-kb band and the plasmid with the distal E2 BS gives rise to a 2.9-kb band.
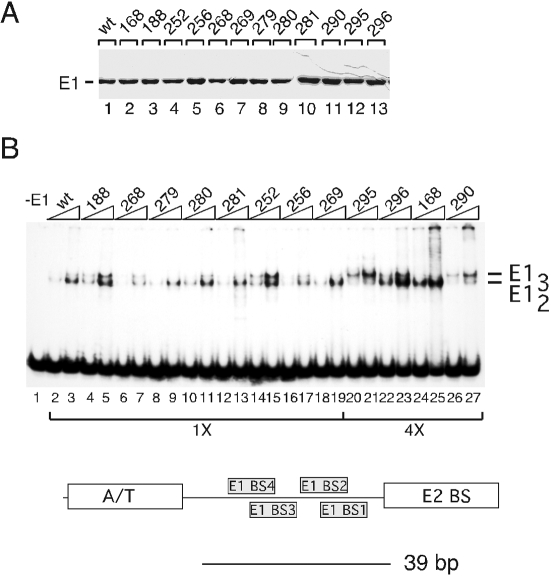
(A) Purification of mutant E1 proteins from E. coli. The mutant E1 proteins were expressed in E. coli and purified using established procedures. wt E1 (lane 1) and the mutant proteins were analyzed by SDS-PAGE and stained with Coomassie brilliant blue. (B) DNA binding activity of mutant E1 proteins. The 12 mutant E1 proteins were tested for sequence-specific ori binding by EMSA in the presence of ADP using a 39-bp probe centered on the E1 BS. Two quantities, 30 and 60 fmol, of the wt and mutant proteins were used in lanes 1 through 19. For the mutants W295A, I296A, K168A, and G290A, which had reduced DNA binding activity, 120 and 240 fmols of E1 were used (lanes 20 through 27).
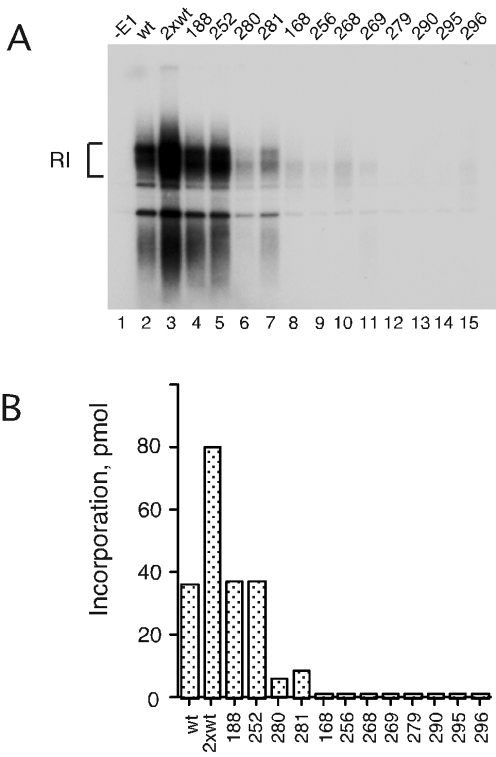
(A) In vitro DNA replication using mutant E1 proteins. The 12 mutant E1 proteins were tested for their abilities to support DNA replication in a cell-free replication system, followed by analysis of the replication products by agarose gel electrophoresis. In lane 1, no E1 was added. In lane 2, 200 ng of wt E1 was added, and in lane 3, 400 ng of wt E1 was added. In lanes 4 through 15, 200 ng of the respective mutant E1 was added. (B) The level of incorporation of [α-32P]dCTP for wt E1 and for each mutant was quantitated and is shown in a bar graph.
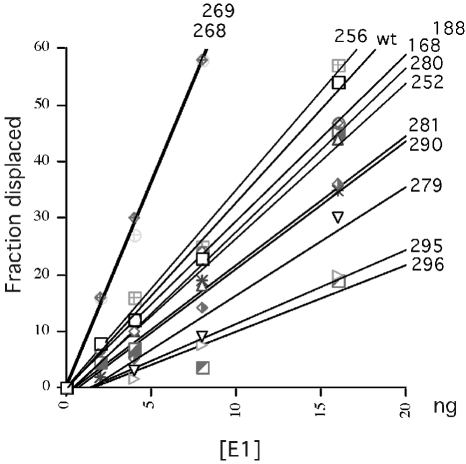
DNA helicase activity of mutant E1 proteins. Three quantities of wt and mutant E1 proteins (8, 15, and 30 fmol) were tested for the ability to displace a labeled oligonucleotide annealed to an M13 template at 37°C. The displaced labeled oligonucleotide was quantitated, and the fraction of input was plotted as a function of E1 levels.
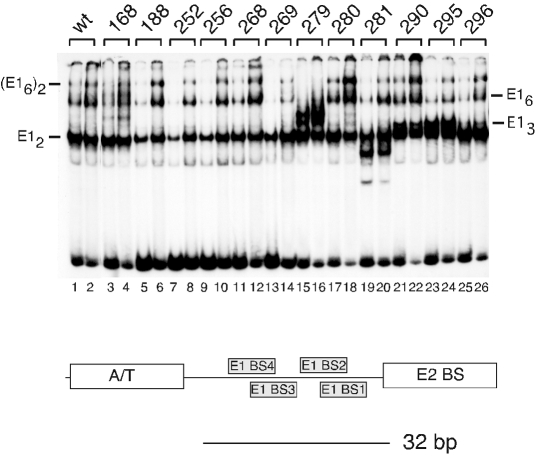
Double hexamer formation by E1 mutants. EMSA was performed using the 32-bp ori probe. Sixty and 120 fmol of wt and mutant E1, respectively, were used in the presence of ATP.
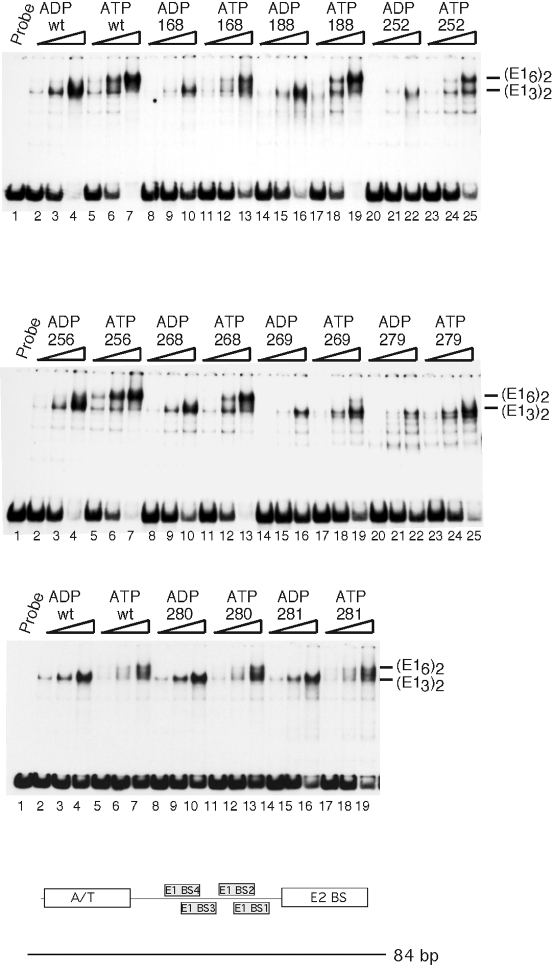
Formation of double trimers and double hexamers by E1 mutants. EMSA was performed using the 84-bp ori probe. Thirty, 60, or 120 fmol of wt and mutant E1 was used in the presence of ATP or ADP as indicated in the figure. Lane 1 in all three panels contained probe alone.
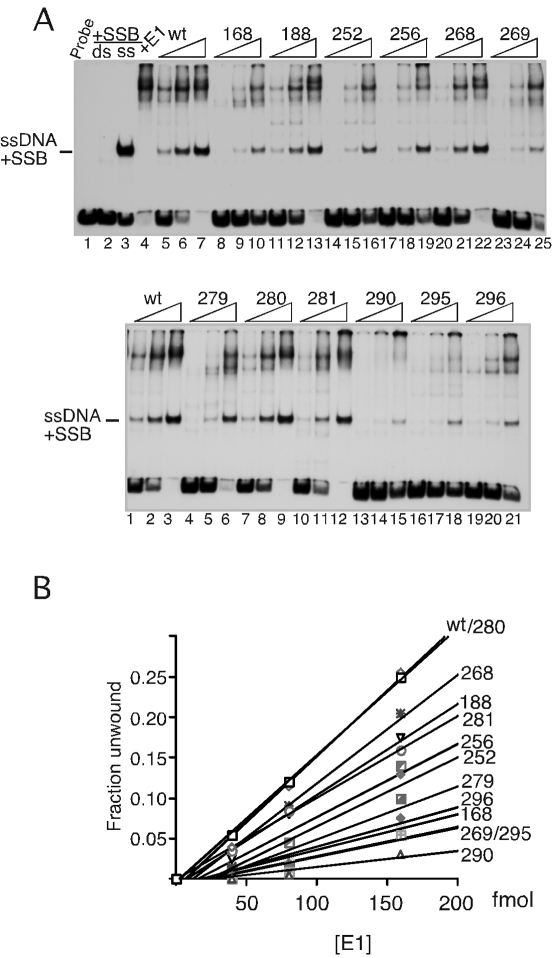
(A) Ori fragment unwinding by E1 mutants. Ori fragment unwinding was carried out using the 84-bp ori probe. In lane 1, top panel, probe alone was added. In lanes 2 and 3, top panel, E. coli SSB was added to double-stranded probe (lane 2) or denatured probe (lane 3). In lane 4, top panel, wt E1 was added in the absence of SSB. In lanes 5 through 25, top panel, and lanes 1 through 21, bottom panel, 40, 80, and 160 fmol of wt E1 and the respective mutants were used. (B) The unwinding reactions in panel A were quantitated, and the fraction of single-stranded DNA generated was plotted as a function of E1 concentration for each mutant.
Similar articles
-
Characterization of the DNA-binding domain of the bovine papillomavirus replication initiator E1.J Virol. 1998 Apr;72(4):2567-76. doi: 10.1128/JVI.72.4.2567-2576.1998. J Virol. 1998. PMID: 9525573 Free PMC article.
-
Two patches of amino acids on the E2 DNA binding domain define the surface for interaction with E1.J Virol. 2000 Feb;74(3):1506-12. doi: 10.1128/jvi.74.3.1506-1512.2000. J Virol. 2000. PMID: 10627562 Free PMC article.
-
Binding of the E1 and E2 proteins to the origin of replication of bovine papillomavirus.J Virol. 1997 Apr;71(4):2887-96. doi: 10.1128/JVI.71.4.2887-2896.1997. J Virol. 1997. PMID: 9060646 Free PMC article.
-
Papillomavirus E1 proteins: form, function, and features.Virus Genes. 2002 Jun;24(3):275-90. doi: 10.1023/a:1015336817836. Virus Genes. 2002. PMID: 12086149 Review.
-
Papillomavirus DNA replication.Intervirology. 1994;37(3-4):150-8. doi: 10.1159/000150373. Intervirology. 1994. PMID: 7843926 Review. No abstract available.
Cited by
-
Mechanisms of Viral DNA Replication of Human Papillomavirus: E2 Protein-Dependent Recruitment of E1 DNA Helicase to the Origin of DNA Replication.Int J Mol Sci. 2025 May 2;26(9):4333. doi: 10.3390/ijms26094333. Int J Mol Sci. 2025. PMID: 40362569 Free PMC article.
-
The papillomavirus E2 proteins.Virology. 2013 Oct;445(1-2):57-79. doi: 10.1016/j.virol.2013.06.006. Epub 2013 Jul 10. Virology. 2013. PMID: 23849793 Free PMC article. Review.
-
ATP-dependent minor groove recognition of TA base pairs is required for template melting by the E1 initiator protein.J Virol. 2007 Apr;81(7):3293-302. doi: 10.1128/JVI.02432-06. Epub 2007 Jan 3. J Virol. 2007. PMID: 17202221 Free PMC article.
-
The E1 proteins.Virology. 2013 Oct;445(1-2):35-56. doi: 10.1016/j.virol.2013.07.020. Epub 2013 Sep 10. Virology. 2013. PMID: 24029589 Free PMC article. Review.
-
'Modulation of the enzymatic activities of replicative helicase (DnaB) by interaction with Hp0897: a possible mechanism for helicase loading in Helicobacter pylori'.Nucleic Acids Res. 2016 Apr 20;44(7):3288-303. doi: 10.1093/nar/gkw148. Epub 2016 Mar 21. Nucleic Acids Res. 2016. PMID: 27001508 Free PMC article.
References
-
- Bullock, P. A. 1997. The initiation of simian virus 40 DNA replication in vitro. Crit. Rev. Biochem. Mol. Biol. 32:503-568. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
