

The mu opioid agonist morphine modulates potentiation of capsaicin-evoked TRPV1 responses through a cyclic AMP-dependent protein kinase A pathway
- PMID: 16842630
- PMCID: PMC1553434
- DOI: 10.1186/1744-8069-2-22
The mu opioid agonist morphine modulates potentiation of capsaicin-evoked TRPV1 responses through a cyclic AMP-dependent protein kinase A pathway
Abstract
Background: The vanilloid receptor 1 (TRPV1) is critical in the development of inflammatory hyperalgesia. Several receptors including G-protein coupled prostaglandin receptors have been reported to functionally interact with the TRPV1 through a cAMP-dependent protein kinase A (PKA) pathway to potentiate TRPV1-mediated capsaicin responses. Such regulation may have significance in inflammatory pain. However, few functional receptor interactions that inhibit PKA-mediated potentiation of TRPV1 responses have been described.
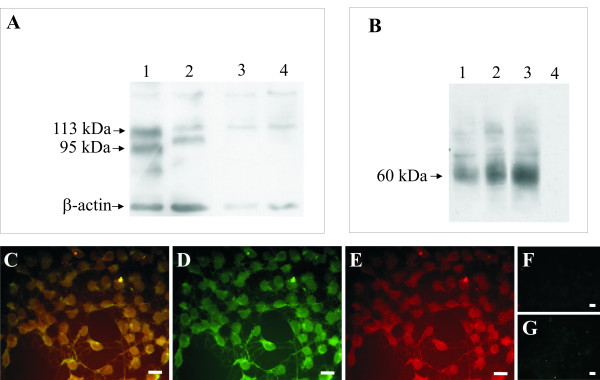
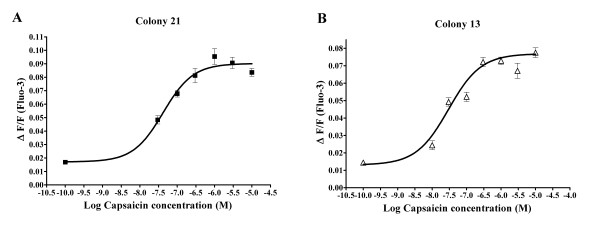
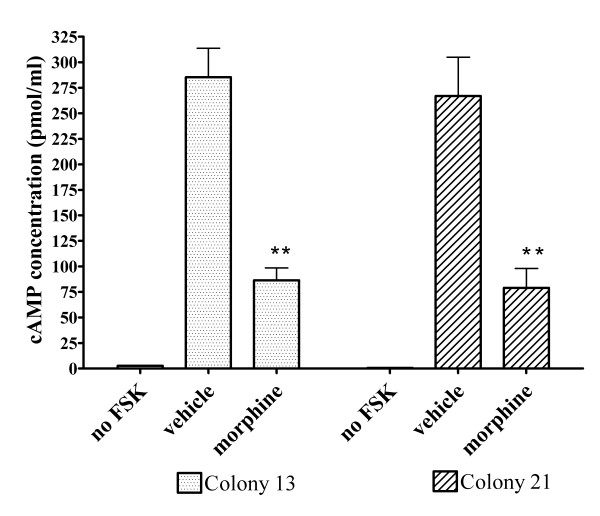
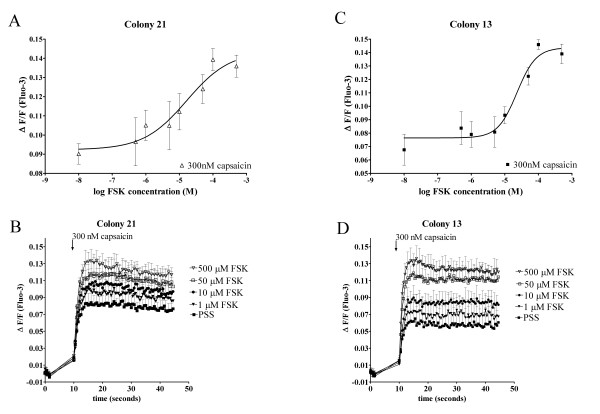
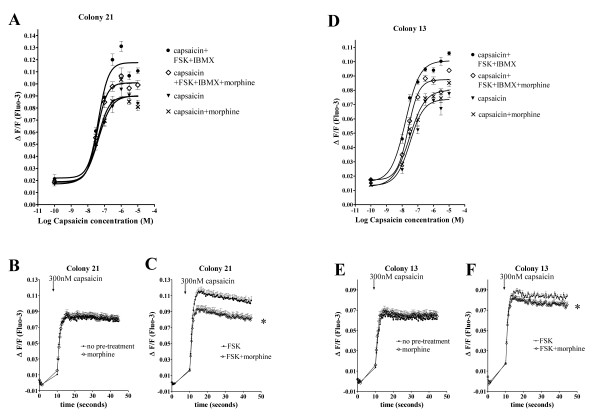
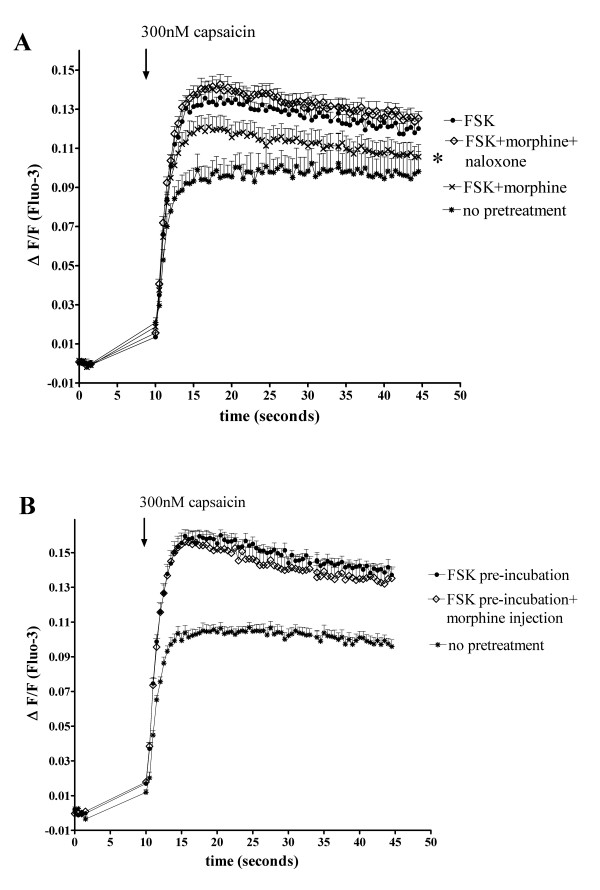
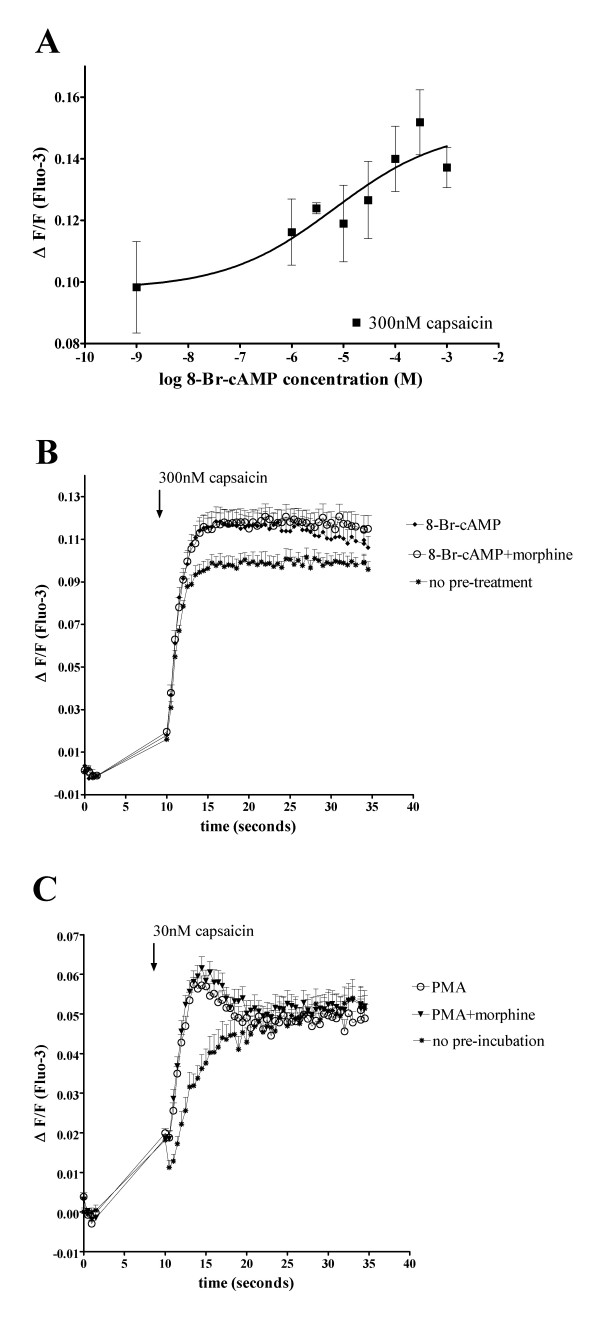
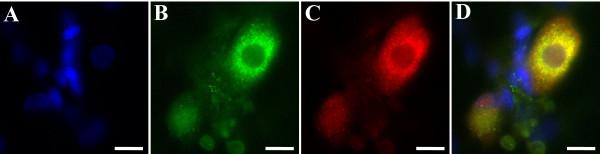
Results: In the present studies we investigated the hypothesis that the mu opioid receptor (MOP) agonist morphine can modulate forskolin-potentiated capsaicin responses through a cAMP-dependent PKA pathway. HEK293 cells were stably transfected with TRPV1 and MOP, and calcium (Ca2+) responses to injection of the TRPV1 agonist capsaicin were monitored in Fluo-3-loaded cells. Pre-treatment with morphine did not inhibit unpotentiated capsaicin-induced Ca2+ responses but significantly altered capsaicin responses potentiated by forskolin. TRPV1-mediated Ca2+ responses potentiated by the direct PKA activator 8-Br-cAMP and the PKC activator Phorbol-12-myristate-13-acetatewere not modulated by morphine. Immunohistochemical studies confirmed that the TRPV1 and MOP are co-expressed on cultured Dorsal Root Ganglion neurones, pointing towards the existence of a functional relationship between the G-protein coupled MOP and nociceptive TRPV1.
Conclusion: The results presented here indicate that the opioid receptor agonist morphine acts via inhibition of adenylate cyclase to inhibit PKA-potentiated TRPV1 responses. Targeting of peripheral opioid receptors may therefore have therapeutic potential as an intervention to prevent potentiation of TRPV1 responses through the PKA pathway in inflammation.
Figures
References
-
- Pomonis JD, Harrison JE, Mark L, Bristol DR, Valenzano KJ, Walker K. N-(4-Tertiarybutylphenyl)-4-(3-cholorphyridin-2-yl)tetrahydropyrazine -1(2H)-carbox-amide (BCTC), a novel, orally effective vanilloid receptor 1 antagonist with analgesic properties: II. in vivo characterization in rat models of inflammatory and neuropathic pain. J Pharmacol Exp Ther. 2003;306:387–393. doi: 10.1124/jpet.102.046268. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
