

CheA Kinase of bacterial chemotaxis: chemical mapping of four essential docking sites
- PMID: 16846213
- PMCID: PMC2904553
- DOI: 10.1021/bi060580y
CheA Kinase of bacterial chemotaxis: chemical mapping of four essential docking sites
Abstract
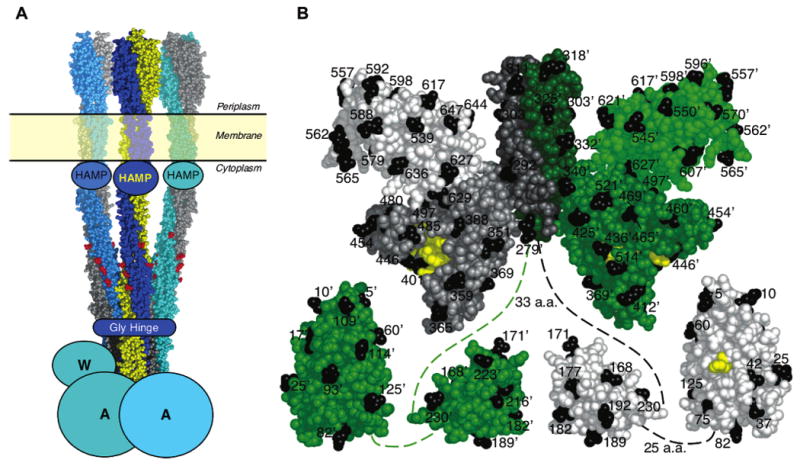
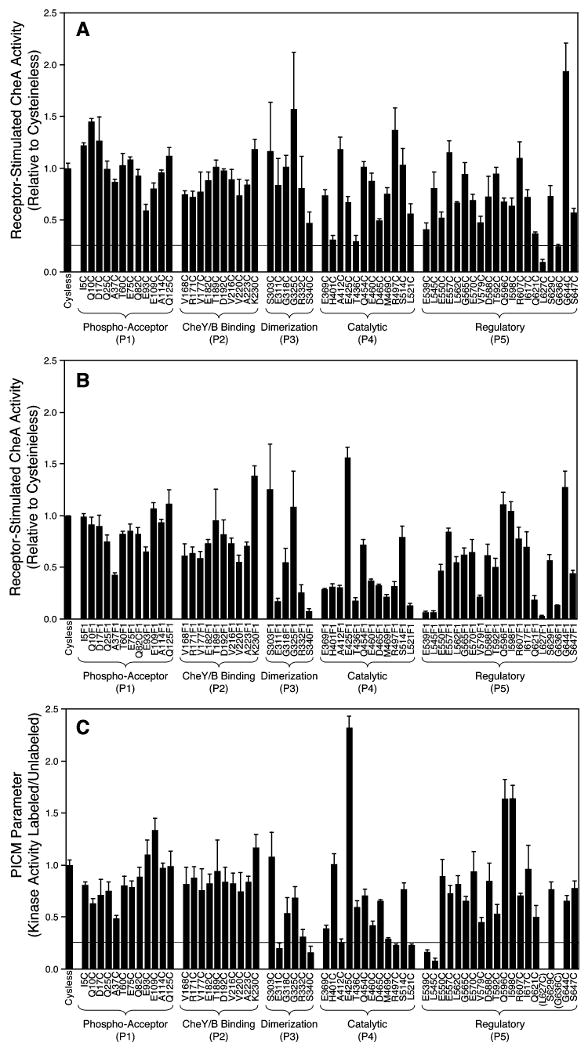
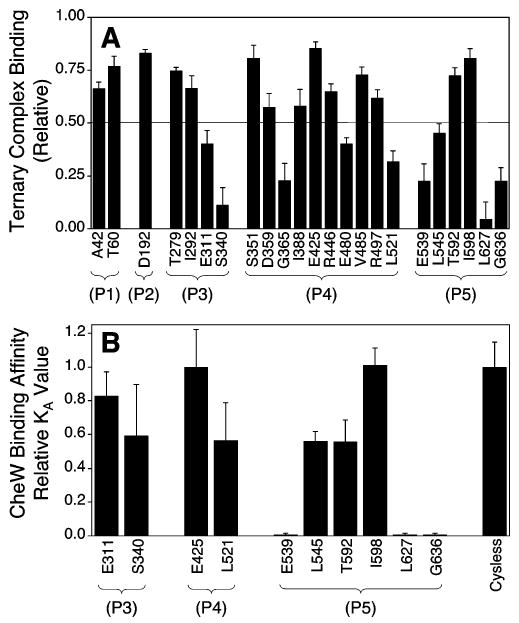
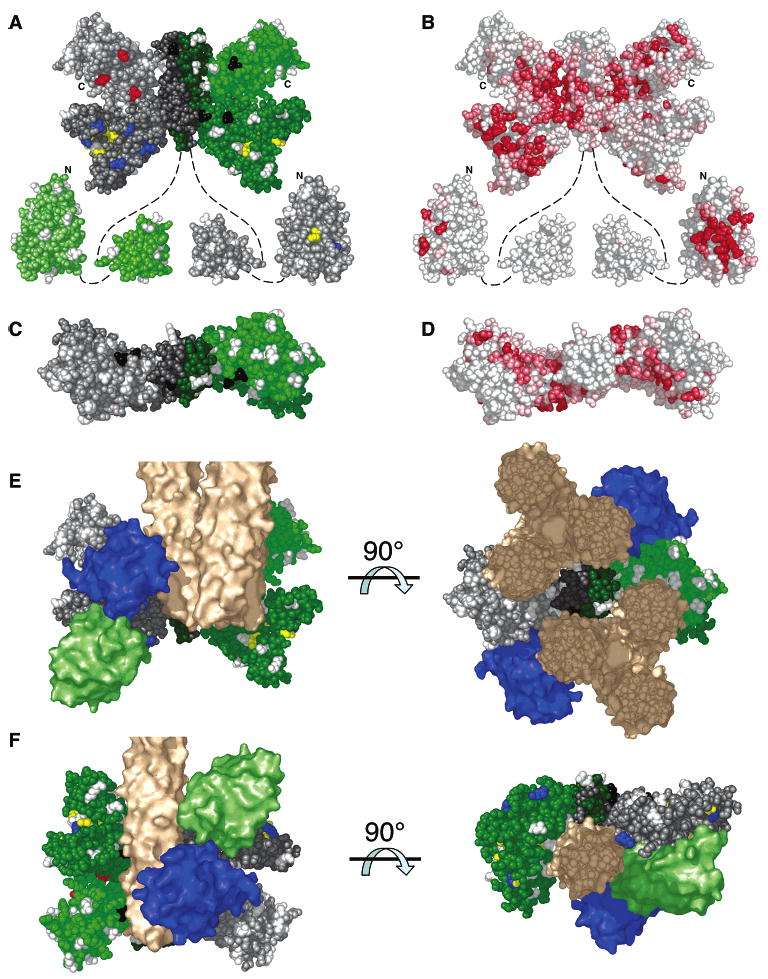
The chemotaxis pathway of Escherichia coli and Salmonella typhimurium is the paradigm for the ubiquitous class of 2-component signaling pathways in prokaryotic organisms. Chemosensing begins with the binding of a chemical attractant to a transmembrane receptor on the cell surface. The resulting transmembrane signal regulates a cytoplasmic, multiprotein signaling complex that controls cellular swimming behavior by generating a diffusible phosphoprotein. The minimal functional unit of this signaling complex, termed the core complex, consists of the transmembrane receptor, the coupling protein CheW, and the histidine kinase CheA. Though the structures of individual components are largely known and the core complex can be functionally reconstituted, the architecture of the assembled core complex has remained elusive. To probe this architecture, the present study has utilized an enhanced version of the protein-interactions-by-cysteine-modification method (PICM-beta) to map out docking surfaces on CheA essential for kinase activity and for core complex assembly. The approach employed a library of 70 single, engineered cysteine residues, scattered uniformly over the surfaces of the five CheA domains in a cysteine-free CheA background. These surface Cys residues were further modified by the sulfhydryl-specific alkylating agent, 5-fluorescein-maleimide (5FM). The functional effects of individual Cys and 5FM-Cys surface modifications were measured by kinase assays of CheA activity in both the free and core complex-associated states, and by direct binding assays of CheA associations with CheW and the receptor. The results define (i) two mutual docking surfaces on the CheA substrate and catalytic domains essential for the association of these domains during autophosphorylation, (ii) a docking surface on the CheA regulatory domain essential for CheW binding, and (iii) a large docking surface encompassing regions of the CheA dimerization, catalytic, and regulatory domains proposed to bind the receptor. To test the generality of these findings, a CheA sequence alignment was analyzed, revealing that the newly identified docking surfaces are highly conserved among CheA homologues. These results strongly suggest that the same docking sites are widely utilized in prokaryotic sensory pathways. Finally, the results provide new structural constraints allowing the development of improved models for core complex architecture.
Figures
Similar articles
-
Defining a key receptor-CheA kinase contact and elucidating its function in the membrane-bound bacterial chemosensory array: a disulfide mapping and TAM-IDS Study.Biochemistry. 2013 Jun 4;52(22):3866-80. doi: 10.1021/bi400385c. Epub 2013 May 23. Biochemistry. 2013. PMID: 23668882 Free PMC article.
-
Structure of the ternary complex formed by a chemotaxis receptor signaling domain, the CheA histidine kinase, and the coupling protein CheW as determined by pulsed dipolar ESR spectroscopy.Biochemistry. 2010 May 11;49(18):3824-41. doi: 10.1021/bi100055m. Biochemistry. 2010. PMID: 20355710 Free PMC article.
-
Cysteine-scanning analysis of the chemoreceptor-coupling domain of the Escherichia coli chemotaxis signaling kinase CheA.J Bacteriol. 2006 Jun;188(12):4321-30. doi: 10.1128/JB.00274-06. J Bacteriol. 2006. PMID: 16740938 Free PMC article.
-
Bacterial chemotaxis coupling protein: Structure, function and diversity.Microbiol Res. 2019 Feb;219:40-48. doi: 10.1016/j.micres.2018.11.001. Epub 2018 Nov 6. Microbiol Res. 2019. PMID: 30642465 Review.
-
pH sensing in bacterial chemotaxis.Novartis Found Symp. 1999;221:38-50; discussions 50-4. doi: 10.1002/9780470515631.ch4. Novartis Found Symp. 1999. PMID: 10207912 Review.
Cited by
-
Structural biology by mass spectrometry: mapping protein interaction surfaces of membrane receptor complexes with ICAT.J Mol Biol. 2011 Jun 17;409(4):481-2. doi: 10.1016/j.jmb.2011.04.030. Epub 2011 Apr 16. J Mol Biol. 2011. PMID: 21515283 Free PMC article. No abstract available.
-
CryoEM and computer simulations reveal a novel kinase conformational switch in bacterial chemotaxis signaling.Elife. 2015 Nov 19;4:e08419. doi: 10.7554/eLife.08419. Elife. 2015. PMID: 26583751 Free PMC article.
-
The core signaling proteins of bacterial chemotaxis assemble to form an ultrastable complex.Biochemistry. 2009 Jul 28;48(29):6975-87. doi: 10.1021/bi900641c. Biochemistry. 2009. PMID: 19456111 Free PMC article.
-
Protein footprinting in a complex milieu: identifying the interaction surfaces of the chemotaxis adaptor protein CheW.J Mol Biol. 2011 Jun 17;409(4):483-95. doi: 10.1016/j.jmb.2011.03.040. Epub 2011 Apr 2. J Mol Biol. 2011. PMID: 21463637 Free PMC article.
-
Structural signatures of Escherichia coli chemoreceptor signaling states revealed by cellular crosslinking.Proc Natl Acad Sci U S A. 2022 Jul 12;119(28):e2204161119. doi: 10.1073/pnas.2204161119. Epub 2022 Jul 5. Proc Natl Acad Sci U S A. 2022. PMID: 35787052 Free PMC article.
References
-
- Parkinson JS, Ames P, Studdert CA. Collaborative signaling by bacterial chemoreceptors. Curr Opin Microbiol. 2005;8:116–121. - PubMed
-
- Wadhams GH, Armitage JP. Making sense of it all: bacterial chemotaxis. Nat Rev Mol Cell Biol. 2004;5:1024–1037. - PubMed
-
- Wolanin PM, Stock JB. Bacterial chemosensing: cooperative molecular logic. Curr Biol. 2004;14:R486–487. - PubMed
-
- Bourret RB, Stock AM. Molecular information processing: lessons from bacterial chemotaxis. J Biol Chem. 2002;277:9625–9628. - PubMed
-
- Spudich JL, Luecke H. Sensory rhodopsin II: functional insights from structure. Curr Opin Struct Biol. 2002;12:540–546. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
