

Visual cycle: Dependence of retinol production and removal on photoproduct decay and cell morphology
- PMID: 16847097
- PMCID: PMC2151530
- DOI: 10.1085/jgp.200609557
Visual cycle: Dependence of retinol production and removal on photoproduct decay and cell morphology
Abstract
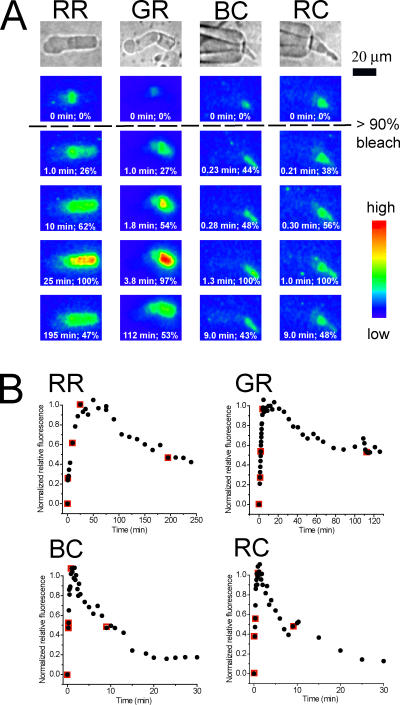
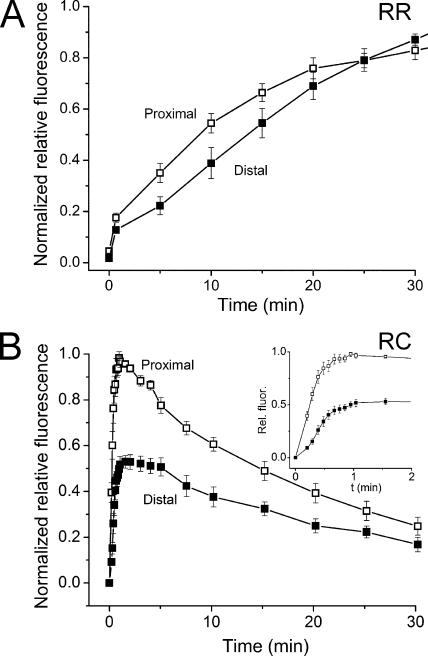
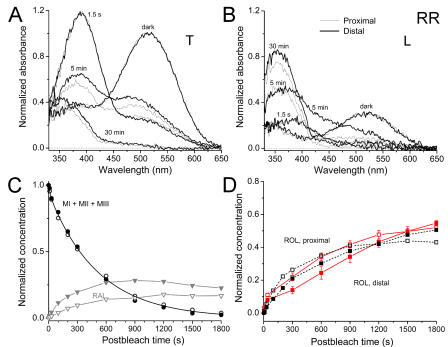
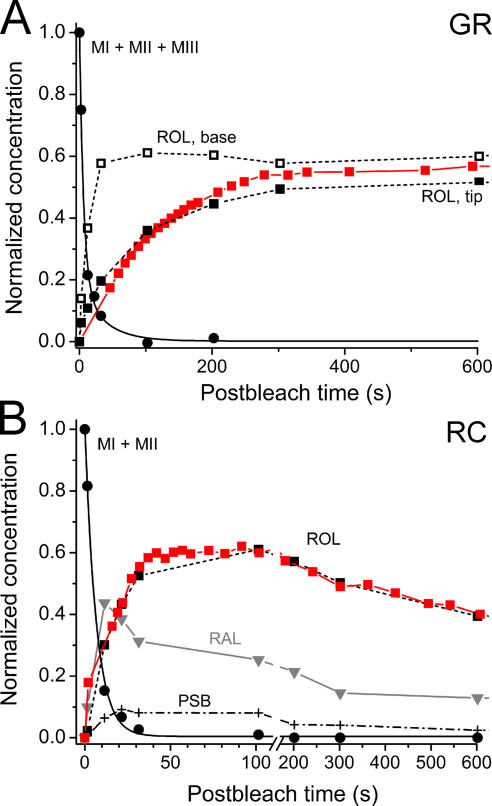
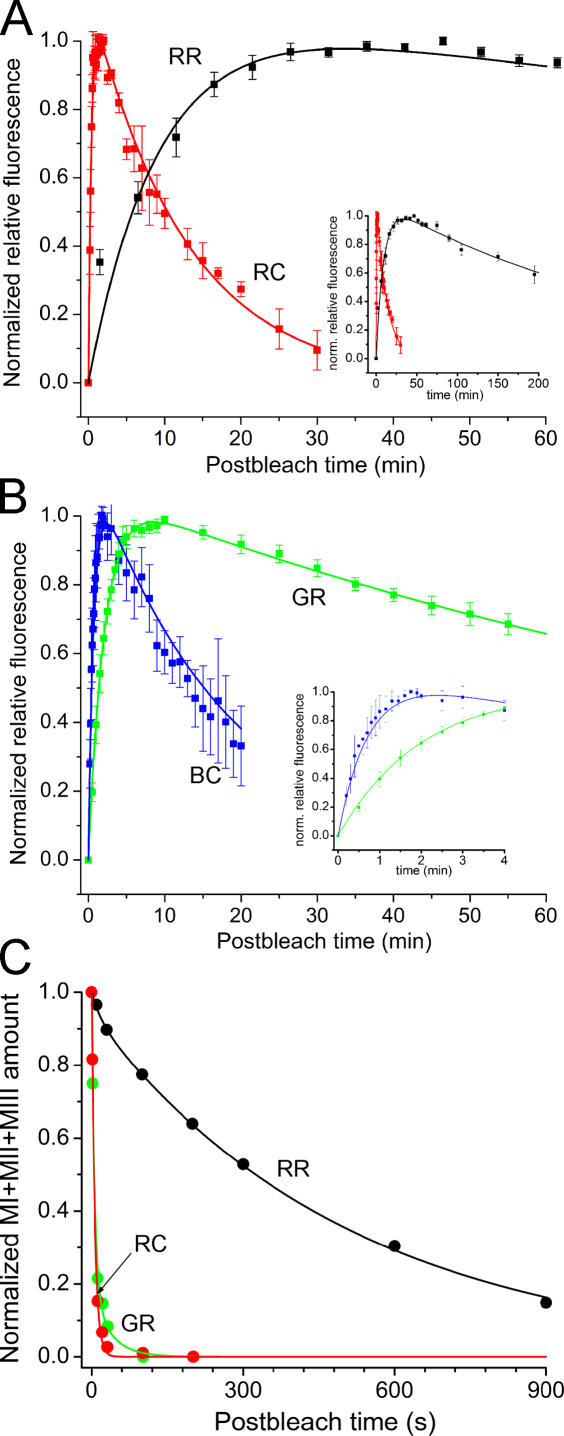
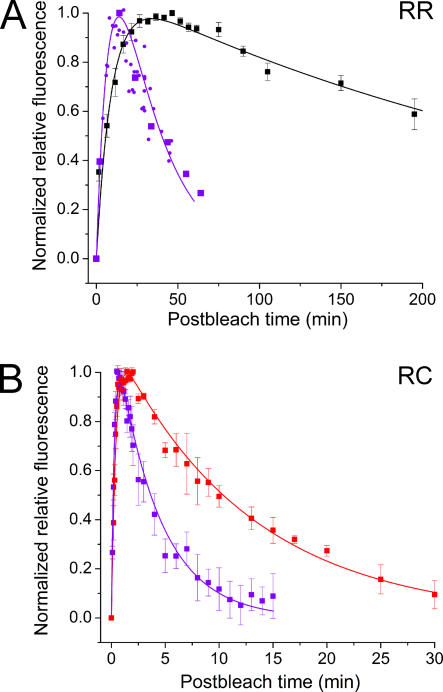
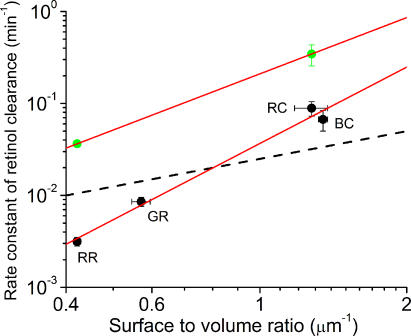
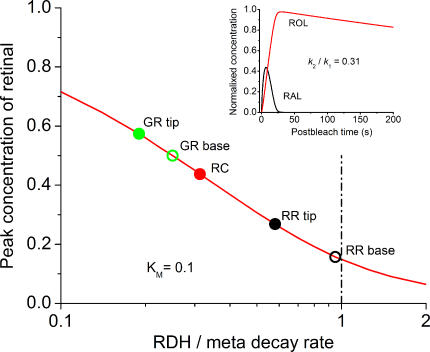
The visual cycle is a chain of biochemical reactions that regenerate visual pigment following exposure to light. Initial steps, the liberation of all-trans retinal and its reduction to all-trans retinol by retinol dehydrogenase (RDH), take place in photoreceptors. We performed comparative microspectrophotometric and microfluorometric measurements on a variety of rod and cone photoreceptors isolated from salamander retinae to correlate the rates of photoproduct decay and retinol production. Metapigment decay rate was spatially uniform within outer segments and 50-70 times faster in the cells that contained cone-type pigment (SWS2 and M/LWS) compared to cells with rod-type pigment (RH1). Retinol production rate was strongly position dependent, fastest at the base of outer segments. Retinol production rate was 10-40 times faster in cones with cone pigments (SWS2 and M/LWS) than in the basal OS of rods containing rod pigment (RH1). Production rate was approximately five times faster in rods containing cone pigment (SWS2) than the rate in basal OS of rods containing the rod pigment (RH1). We show that retinol production is defined either by metapigment decay rate or RDH reaction rate, depending on cell type or outer segment region, whereas retinol removal is defined by the surface-to-volume ratio of the outer segment and the availability of retinoid binding protein (IRBP). The more rapid rates of retinol production in cones compared to rods are consistent with the more rapid operation of the visual cycle in these cells.
Figures
Comment in
-
Timing is everything: Direct measurement of retinol production in cones and rods.J Gen Physiol. 2006 Aug;128(2):147-8. doi: 10.1085/jgp.200609609. Epub 2006 Jul 17. J Gen Physiol. 2006. PMID: 16847095 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
