

Understanding the molecular basis of the interaction between NDPK-A and AMPK alpha 1
- PMID: 16847342
- PMCID: PMC1592779
- DOI: 10.1128/MCB.00315-06
Understanding the molecular basis of the interaction between NDPK-A and AMPK alpha 1
Retraction in
-
Understanding the molecular basis of the interaction between NDPK-A and AMPK alpha1.Mol Cell Biol. 2008 Sep;28(18):5827. doi: 10.1128/MCB.00859-08. Mol Cell Biol. 2008. PMID: 18772271 Free PMC article. No abstract available.
Abstract
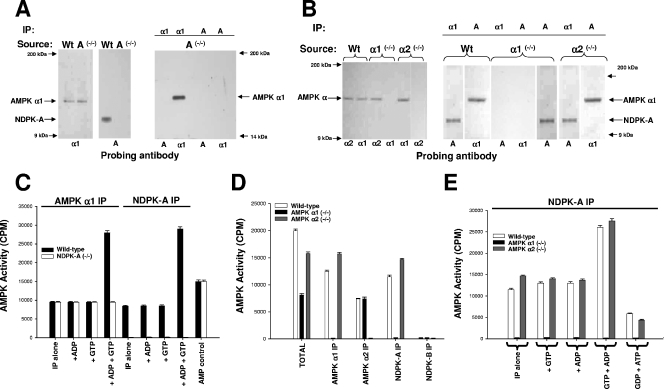
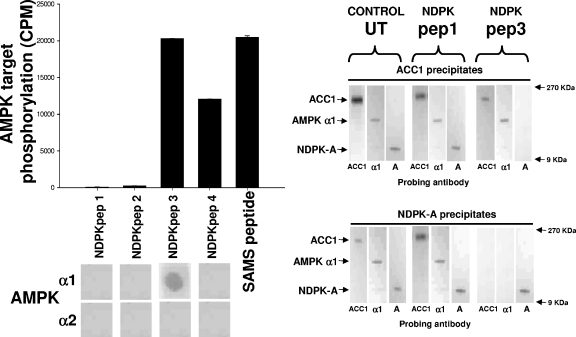
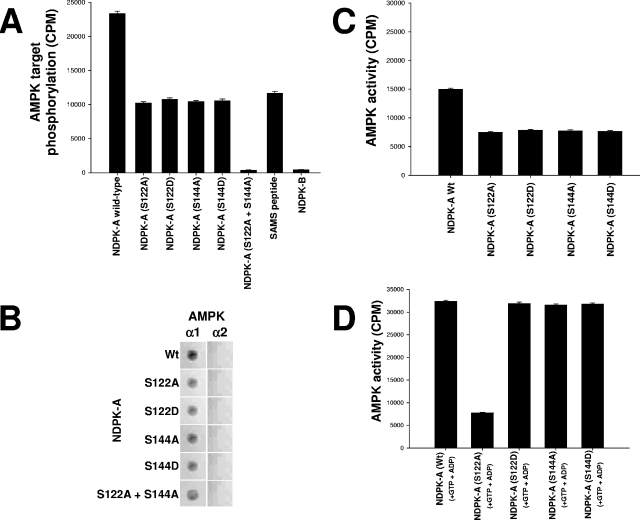
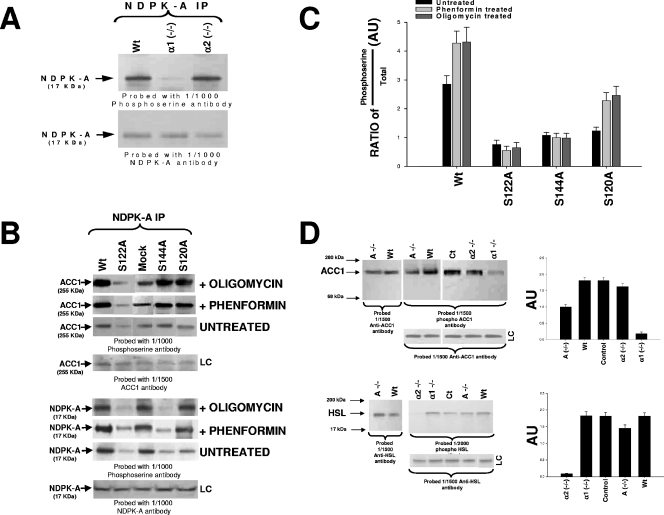
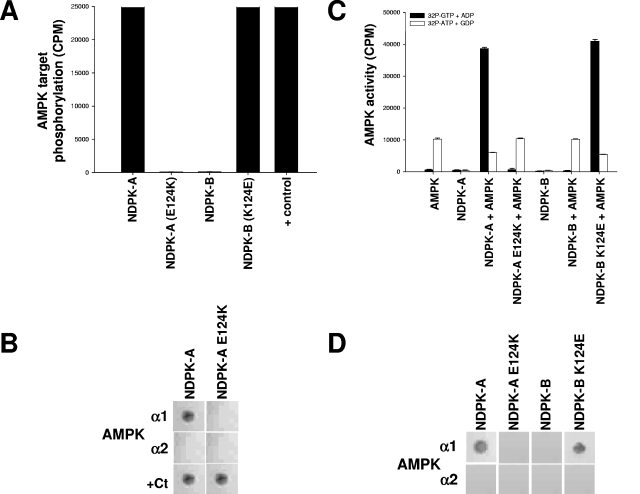
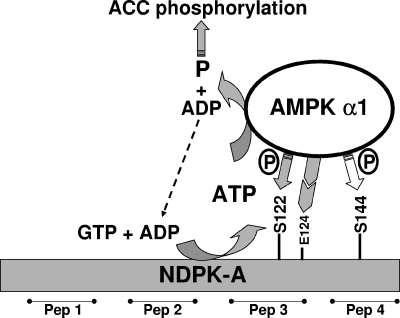
Nucleoside diphosphate kinase (NDPK) (nm23/awd) belongs to a multifunctional family of highly conserved proteins (approximately 16 to 20 kDa) including two well-characterized isoforms (NDPK-A and -B). NDPK catalyzes the conversion of nucleoside diphosphates to nucleoside triphosphates, regulates a diverse array of cellular events, and can act as a protein histidine kinase. AMP-activated protein kinase (AMPK) is a heterotrimeric protein complex that responds to the cellular energy status by switching off ATP-consuming pathways and switching on ATP-generating pathways when ATP is limiting. AMPK was first discovered as an activity that inhibited preparations of acetyl coenzyme A carboxylase 1 (ACC1), a regulator of cellular fatty acid synthesis. We recently reported that NDPK-A (but not NDPK-B) selectively regulates the alpha1 isoform of AMPK independently of the AMP concentration such that the manipulation of NDPK-A nucleotide trans-phosphorylation activity to generate ATP enhanced the activity of AMPK. This regulation occurred irrespective of the surrounding ATP concentration, suggesting that "substrate channeling" was occurring with the shielding of NDPK-generated ATP from the surrounding medium. We speculated that AMPK alpha1 phosphorylated NDPK-A during their interaction, and here, we identify two residues on NDPK-A targeted by AMPK alpha1 in vivo. We find that NDPK-A S122 and S144 are phosphorylated by AMPK alpha1 and that the phosphorylation status of S122, but not S144, determines whether substrate channeling can occur. We report the cellular effects of the S122 mutation on ACC1 phosphorylation and demonstrate that the presence of E124 (absent in NDPK-B) is necessary and sufficient to permit both AMPK alpha1 binding and substrate channeling.
Figures
References
-
- Arnaud-Dabernat, S., P. M. Bourbon, A. Dierich, M. Le Meur, and J. Y. Daniel. 2003. Knockout mice as model systems for studying nm23/NDP kinase gene functions. Application to the nm23-M1 gene. J. Bioenerg. Biomembr. 35:19-30. - PubMed
-
- Biondi, R. M., M. Engel, M. Sauane, C. Welter, O. G. Issinger, L. Jimenez de Asua, and S. Passeron. 1996. Inhibition of nucleoside diphosphate kinase activity by in vitro phosphorylation by protein kinase CK2. Differential phosphorylation of NDP kinases in HeLa cells in culture. FEBS Lett. 399:183-187. - PubMed
-
- Bosnar, M. H., J. De Gunzburg, R. Bago, L. Brecevic, I. Weber, and J. Pavelic. 2004. Subcellular localization of A and B Nm23/NDPK subunits. Exp. Cell Res. 298:275-284. - PubMed
-
- Bradford, M. M. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 72:248-254. - PubMed
-
- Caligo, M. A., G. Cipollini, L. Fiore, S. Calvo, F. Basolo, P. Collecchi, F. Ciardiello, S. Pepe, M. Petrini, and G. Bevilacqua. 1995. NM23 gene expression correlates with cell growth rate and S-phase. Int. J. Cancer 60:837-842. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous