

Molecular nature of Spemann's organizer: the role of the Xenopus homeobox gene goosecoid
- PMID: 1684739
- PMCID: PMC3102583
- DOI: 10.1016/0092-8674(91)90288-a
Molecular nature of Spemann's organizer: the role of the Xenopus homeobox gene goosecoid
Abstract
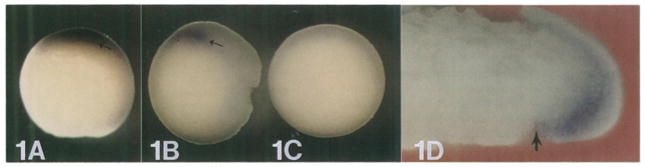
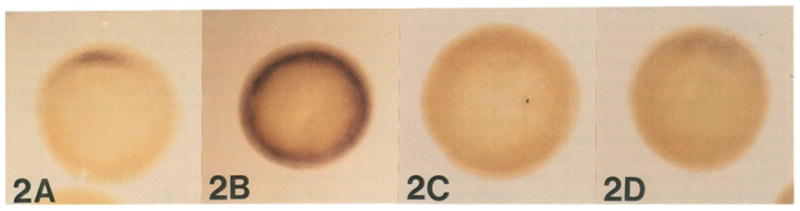
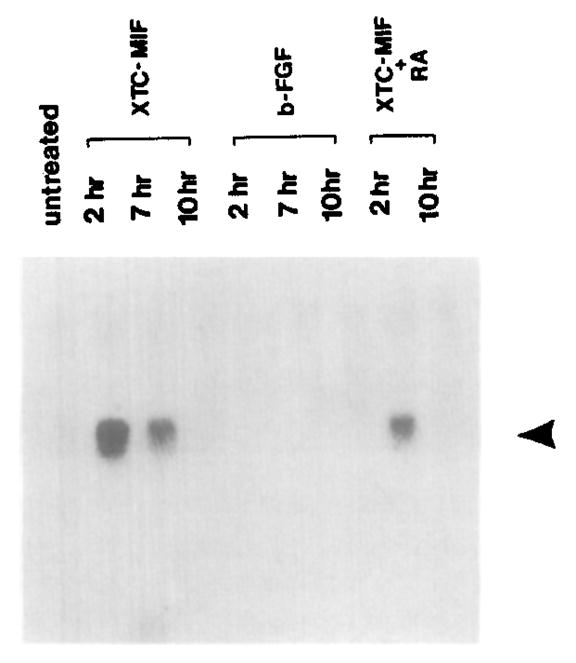
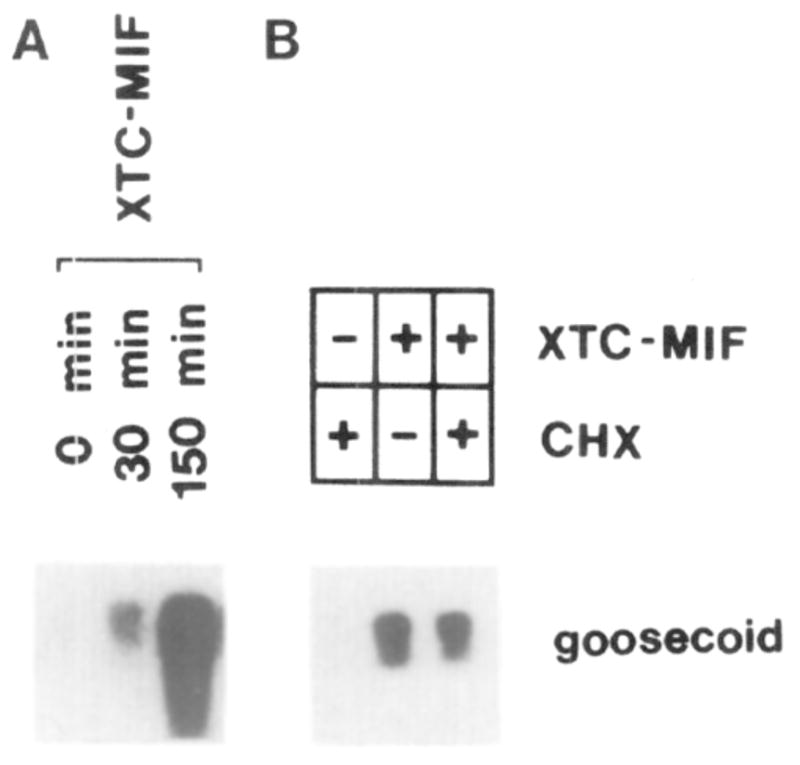
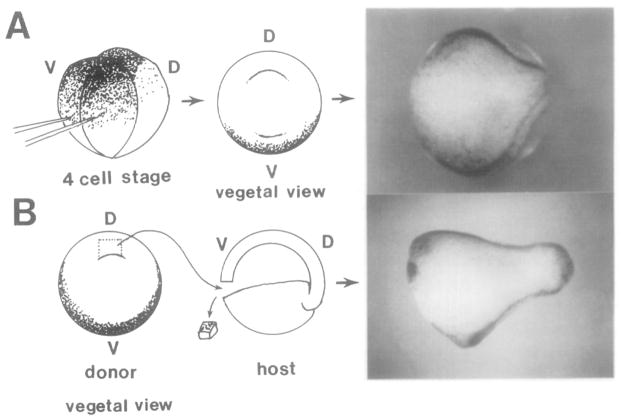
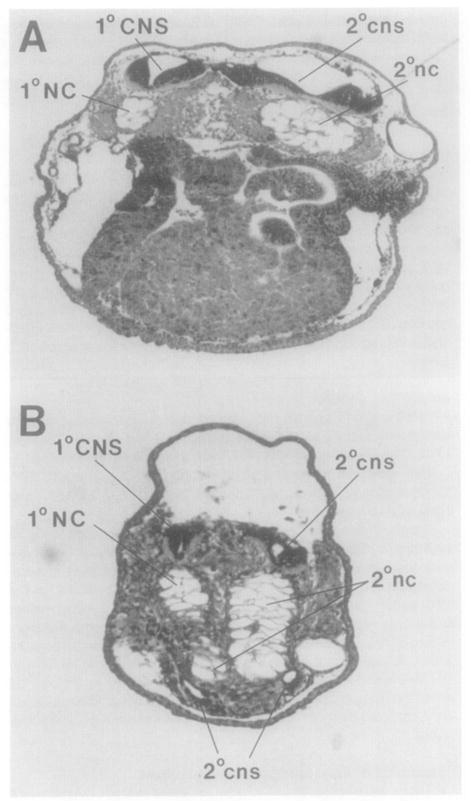
This study analyzes the function of the homeobox gene goosecoid in Xenopus development. First, we find that goosecoid mRNA distribution closely mimics the expected localization of organizer tissue in normal embryos as well as in those treated with LiCl and UV light. Second, goosecoid mRNA accumulation is induced by activin, even in the absence of protein synthesis. It is not affected by bFGF and is repressed by retinoic acid. Lastly, microinjection of goosecoid mRNA into the ventral side of Xenopus embryos, where goosecoid is normally absent, leads to the formation of an additional complete body axis, including head structures and abundant notochordal tissue. The results suggest that the goosecoid homeodomain protein plays a central role in executing Spemann's organizer phenomenon.
Figures

References
-
- Amaya E, Musci TJ, Kirschner MW. Expression of a dominant negative mutant of the FGF receptor disrupts mesoderm formation in Xenopus embryos. Cell. 1991;66:257–270. - PubMed
-
- Blumberg B, Wright CVE, De Robertis EM, Cho KWY. Organizer-specific homeobox genes in Xenopus laevis embryos. Science. 1991;253:194–196. - PubMed
-
- Busa WB, Gimlich RL. Lithium-induced teratogenesis in frog embryos prevented by a polyphosphoinositide cycle intermediate or a diacylglycerol analog. Dev Biol. 1989;132:315–324. - PubMed
-
- Cho KWY, De Robertis EM. Differential activation of Xenopus homeobox genes by mesoderm inducing growth factors and retinoic acid. Genes Dev. 1990;4:1910–1916. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
