

An experimental study into the acousto-mechanical effects of invading the cochlea
- PMID: 16849252
- PMCID: PMC1664639
- DOI: 10.1098/rsif.2006.0117
An experimental study into the acousto-mechanical effects of invading the cochlea
Abstract
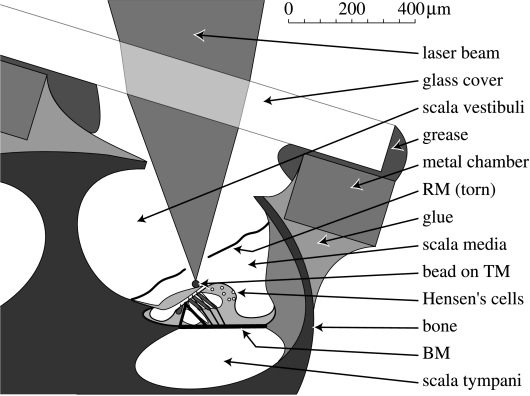
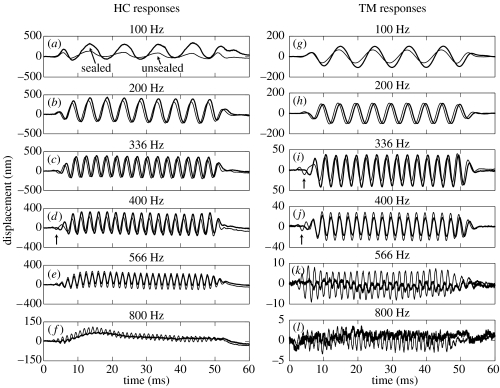
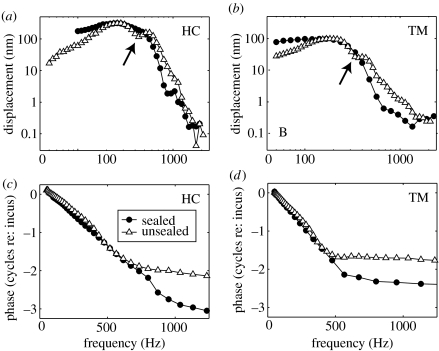
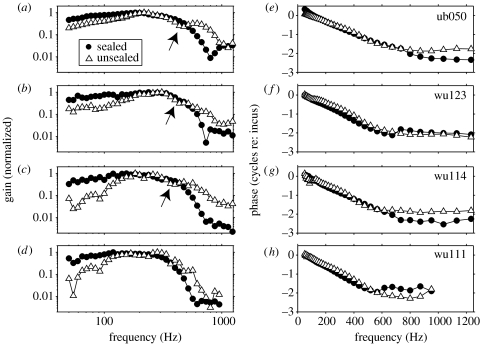
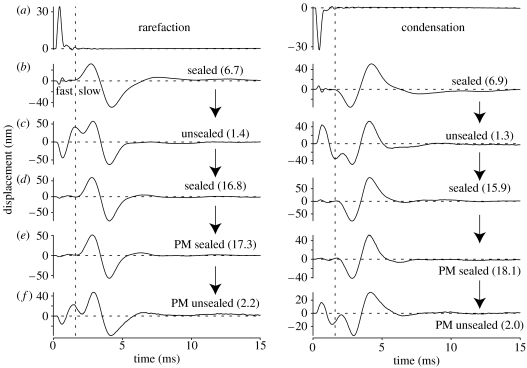
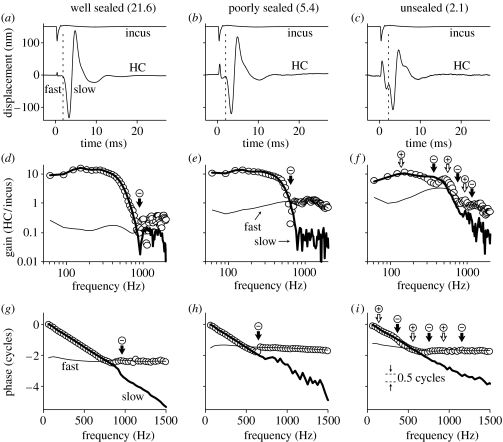
The active and nonlinear mechanical processing of sound that takes place in the mammalian cochlea is fundamental to our sense of hearing. We have investigated the effects of opening the cochlea in order to make experimental observations of this processing. Using an optically transparent window that permits laser interferometric access to the apical turn of the guinea-pig cochlea, we show that the acousto-mechanical transfer functions of the sealed (i.e. near intact) cochlea are considerably simpler than those of the unsealed cochlea. Comparison of our results with those of others suggests that most previous investigations of apical cochlear mechanics have been made under unsealed conditions, and are therefore likely to have misrepresented the filtering of low-frequency sounds in the cochlea. The mechanical filtering that is apparent in the apical turns of sealed cochleae also differs from the filtering seen in individual auditory nerve fibres with similar characteristic frequencies. As previous studies have shown the neural and mechanical tuning of the basal cochlea to be almost identical, we conclude that the strategies used to process low frequency sounds in the apical turns of the cochlea might differ fundamentally from those used to process high frequency sounds in the basal turns.
Figures
References
-
- Allen J.B. Cochlear micromechanics—a physical model of transduction. J. Acoust. Soc. Am. 1980;68:1660–1670. doi:10.1121/1.385198 - DOI - PubMed
-
- Békésy G.v. McGraw-Hill; New York: 1960. Experiments in hearing.
-
- Cooper N.P. Mid-band sensitivity notches in apical cochlear mechanics. In: Hecht-Poinar E, editor. Diversity in auditory mechanics. World Scientific; Singapore: 1996. pp. 298–304.
-
- Cooper N.P. An improved heterodyne laser interferometer for use in studies of cochlear mechanics. J. Neurosci. Methods. 1999;88:93–102. doi:10.1016/S0165-0270(99)00017-5 - DOI - PubMed
-
- Cooper N.P, Dong W. Sound-evoked changes in the baseline position of the cochlear partition at the apex of the guinea-pig cochlea. J. Physiol. 2000;527P:90P.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
