

Maternal gametophytic baseless1 is required for development of the central cell and early endosperm patterning in maize (Zea mays)
- PMID: 16849604
- PMCID: PMC1569813
- DOI: 10.1534/genetics.106.059709
Maternal gametophytic baseless1 is required for development of the central cell and early endosperm patterning in maize (Zea mays)
Abstract
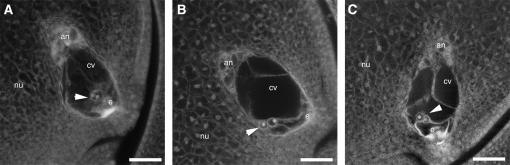
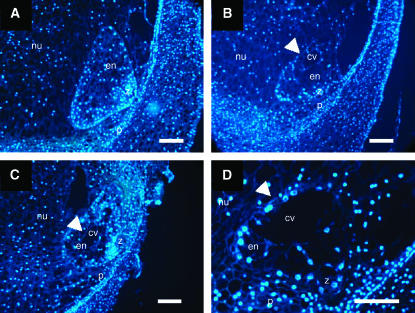
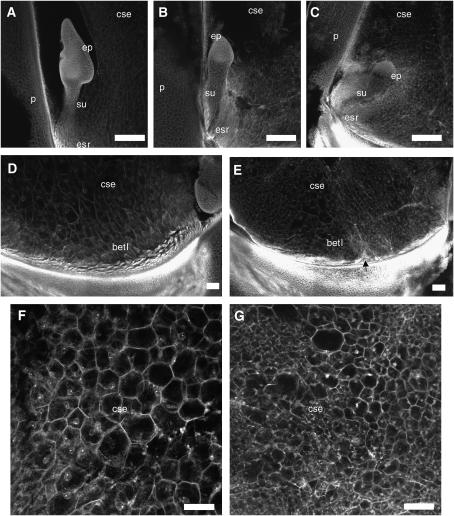
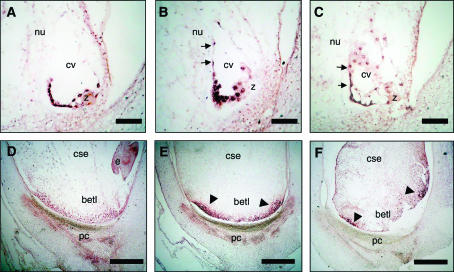
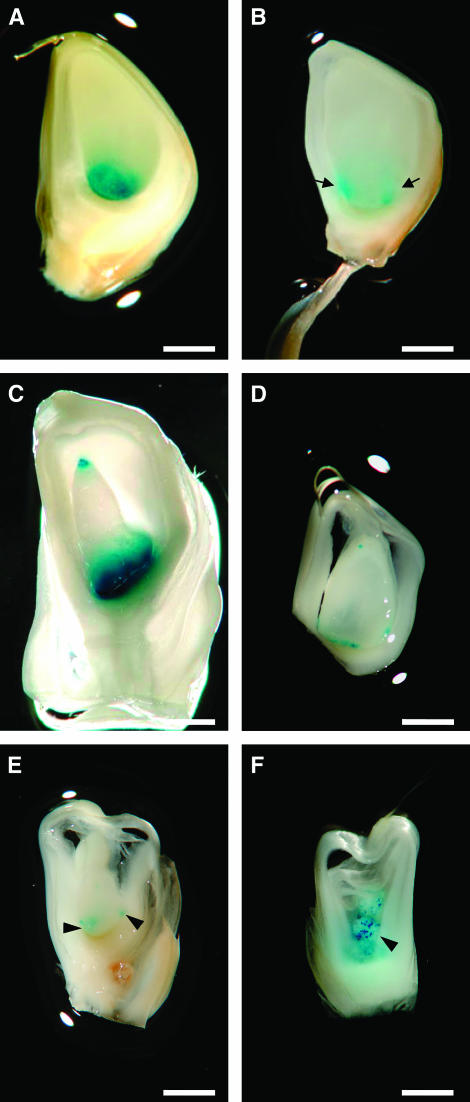
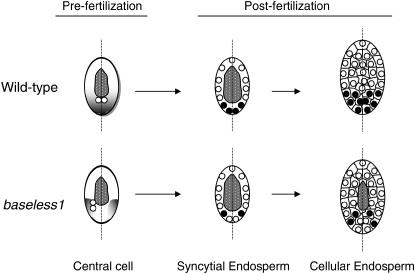
In angiosperms, double fertilization of an egg cell and a central cell with two sperm cells results in the formation of a seed containing a diploid embryo and a triploid endosperm. The extent to which the embryo sac controls postfertilization events in the seed is unknown. The novel gametophytic maternal-effect maize mutation, baseless1 (bsl1) affects central cell development within the embryo sac, frequently by altering the position of the two polar nuclei. Despite this irregularity, fertilization is as efficient as in wild type. The spatial expression of basal endosperm-specific transcripts is altered in free-nuclear and cellular mutant endosperms. At later stages of seed development, bsl1 predominantly affects development of the basal endosperm transfer layer (BETL). When bsl1/+ diploid plants were pollinated by wild-type tetraploid plants, the BETL abnormalities observed in bsl1/bsl1/+/+ tetraploid endosperms were diverse and of variable severity. Moreover, the frequency of kernels with severely perturbed BETL development correlated with the percentage of severely affected bsl1 central cells. Therefore, BSL1 is likely required in the central cell before fertilization for correct BETL patterning to occur. These findings provide new genetic evidence that a maternal gametophytic component is necessary for correct endosperm patterning.
Figures

References
-
- Birchler, J. A., 1993. Dosage analysis of maize endosperm development. Annu. Rev. Genet. 27: 181–204. - PubMed
-
- Brown, R. C., B. E. Lemmon and H. Nguyen, 2003. Events during the first four rounds of mitosis establish three developmental domains in the syncytial endosperm of Arabidopsis thaliana. Protoplasma 222: 167–174. - PubMed
-
- Chang, M. T., and M. G. Neuffer, 1994. Endosperm-embryo interactions in maize. Maydica 39: 9–18.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
