

Yeast-to-hyphal transition triggers formin-dependent Golgi localization to the growing tip in Candida albicans
- PMID: 16855023
- PMCID: PMC1635370
- DOI: 10.1091/mbc.e06-02-0143
Yeast-to-hyphal transition triggers formin-dependent Golgi localization to the growing tip in Candida albicans
Abstract
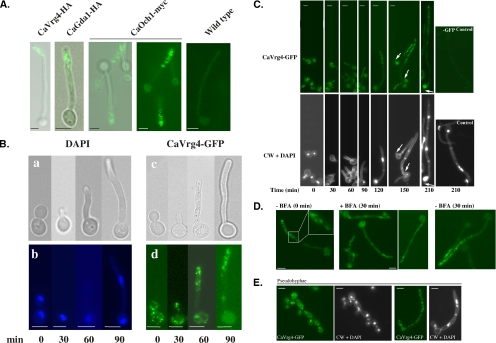
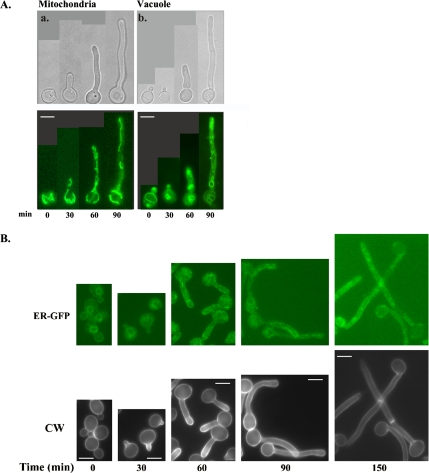
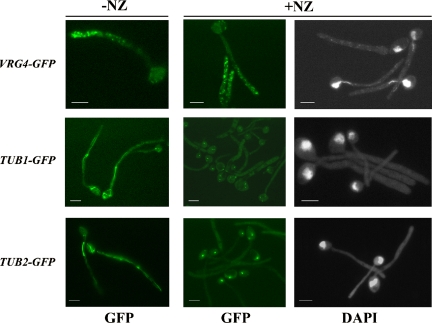
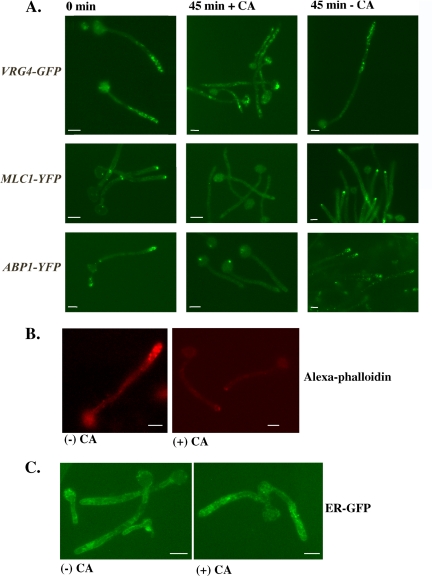
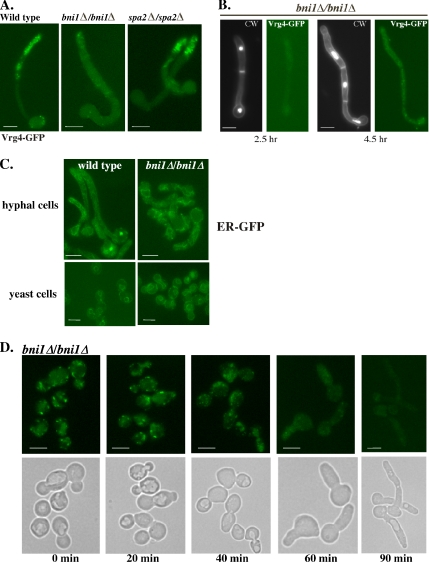
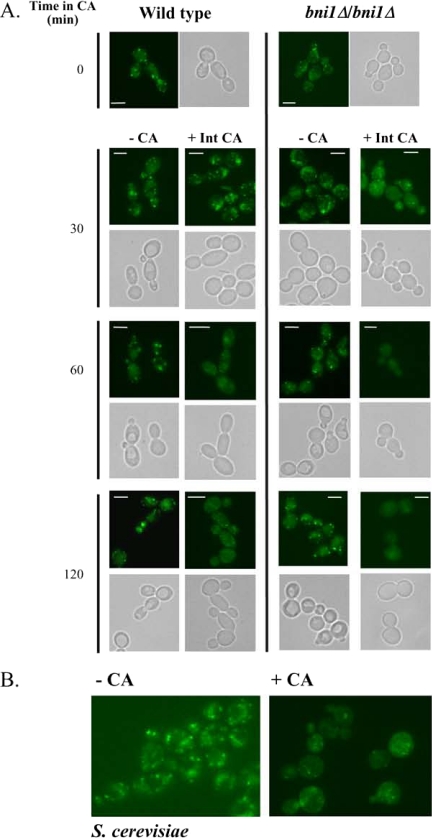
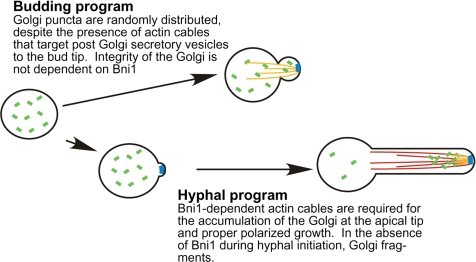
Rapid and long-distance secretion of membrane components is critical for hyphal formation in filamentous fungi, but the mechanisms responsible for polarized trafficking are not well understood. Here, we demonstrate that in Candida albicans, the majority of the Golgi complex is redistributed to the distal region during hyphal formation. Randomly distributed Golgi puncta in yeast cells cluster toward the growing tip during hyphal formation, remain associated with the distal portion of the filament during its extension, and are almost absent from the cell body. This restricted Golgi localization pattern is distinct from other organelles, including the endoplasmic reticulum, vacuole and mitochondria, which remain distributed throughout the cell body and hypha. Hyphal-induced positioning of the Golgi and the maintenance of its structural integrity requires actin cytoskeleton, but not microtubules. Absence of the formin Bni1 causes a hyphal-specific dispersal of the Golgi into a haze of finely dispersed vesicles with a sedimentation density no different from that of normal Golgi. These results demonstrate the existence of a hyphal-specific, Bni1-dependent cue for Golgi integrity and positioning at the distal portion of the hyphal tip, and suggest that filamentous fungi have evolved a novel strategy for polarized secretion, involving a redistribution of the Golgi to the growing tip.
Figures

References
-
- Akashi T., Kanbe T., Tanaka K. The role of the cytoskeleton in the polarized growth of the germ tube in Candida albicans. Microbiology. 1994;140:271–280. - PubMed
-
- Bates S., et al. Outer chain N-glycans are required for cell wall integrity and virulence of Candida albicans. J. Biol. Chem. 2006;281:90–98. - PubMed
-
- Cole L., Orlovich D.A., Ashford A. E. Structure, function and motility of vacuoles in filamentous fungi. Fungal Genet. Biol. 1998;24:86–100. - PubMed
-
- Cormack B. P., Bertram G., Egerton M., Gow N. A., Falkow S., Brown A. J. Yeast-enhanced green fluorescent protein (yEGFP)a reporter of gene expression in Candida albicans. Microbiology. 1997;143:303–311. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
