

Chromosome-wide identification of novel imprinted genes using microarrays and uniparental disomies
- PMID: 16855283
- PMCID: PMC1524921
- DOI: 10.1093/nar/gkl461
Chromosome-wide identification of novel imprinted genes using microarrays and uniparental disomies
Abstract
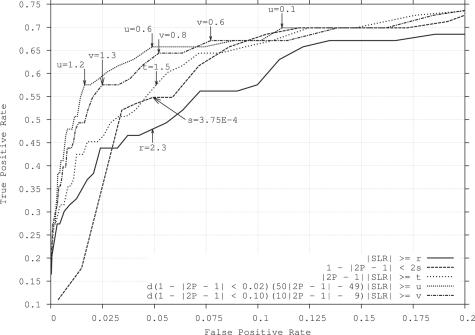
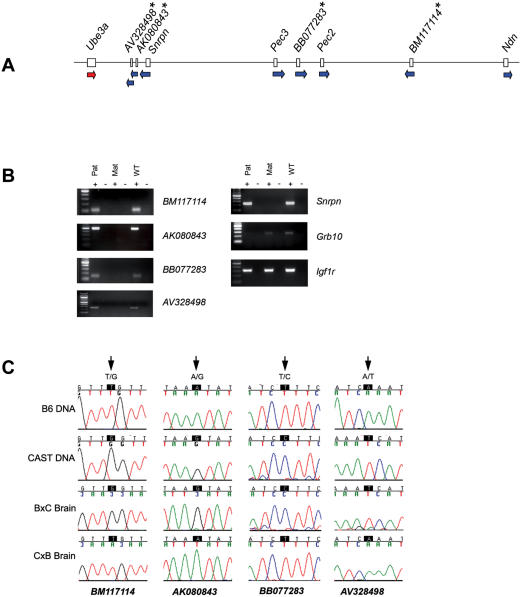
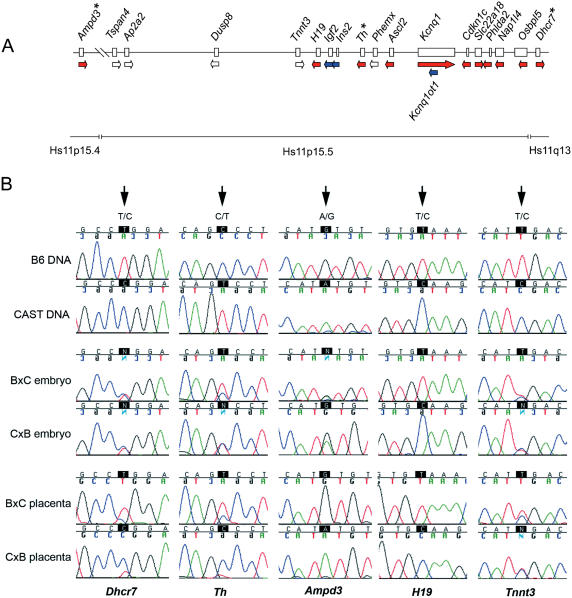
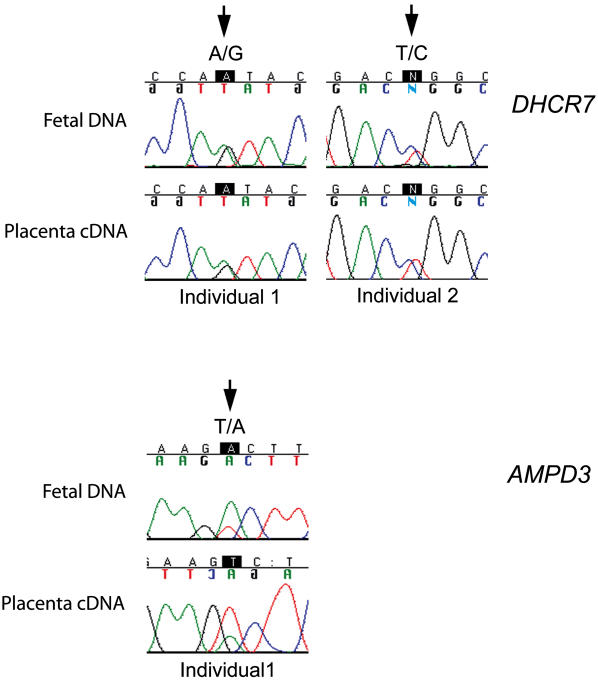
Genomic imprinting refers to a specialized form of epigenetic gene regulation whereby the expression of a given allele is dictated by parental origin. Defining the extent and distribution of imprinting across genomes will be crucial for understanding the roles played by imprinting in normal mammalian growth and development. Using mice carrying uniparental disomies or duplications, microarray screening and stringent bioinformatics, we have developed the first large-scale tissue-specific screen for imprinted gene detection. We quantify the stringency of our methodology and relate it to previous non-tissue-specific large-scale studies. We report the identification in mouse of four brain-specific novel paternally expressed transcripts and an additional three genes that show maternal expression in the placenta. The regions of conserved linkage in the human genome are associated with the Prader-Willi Syndrome (PWS) and Beckwith-Wiedemann Syndrome (BWS) where imprinting is known to be a contributing factor. We conclude that large-scale systematic analyses of this genre are necessary for the full impact of genomic imprinting on mammalian gene expression and phenotype to be elucidated.
Figures
Similar articles
-
An unexpected function of the Prader-Willi syndrome imprinting center in maternal imprinting in mice.PLoS One. 2012;7(4):e34348. doi: 10.1371/journal.pone.0034348. Epub 2012 Apr 4. PLoS One. 2012. PMID: 22496793 Free PMC article.
-
Influence of the Prader-Willi syndrome imprinting center on the DNA methylation landscape in the mouse brain.Epigenetics. 2014 Nov;9(11):1540-56. doi: 10.4161/15592294.2014.969667. Epigenetics. 2014. PMID: 25482058 Free PMC article.
-
Analysis of imprinting in mice with uniparental duplication of proximal chromosomes 7 and 15 by use of a custom oligonucleotide microarray.Mamm Genome. 2004 Mar;15(3):199-209. doi: 10.1007/s00335-003-2322-8. Mamm Genome. 2004. PMID: 15014969
-
Prader-Willi syndrome and Angelman syndrome.Am J Med Genet C Semin Med Genet. 2010 Aug 15;154C(3):365-76. doi: 10.1002/ajmg.c.30273. Am J Med Genet C Semin Med Genet. 2010. PMID: 20803659 Review.
-
Why is there no diploid overdose effect in Prader-Willi syndrome due to uniparental disomy?Acta Genet Med Gemellol (Roma). 1996;45(1-2):179-89. doi: 10.1017/s0001566000001288. Acta Genet Med Gemellol (Roma). 1996. PMID: 8872029 Review.
Cited by
-
Genome-wide mapping of imprinted differentially methylated regions by DNA methylation profiling of human placentas from triploidies.Epigenetics Chromatin. 2011 Jul 13;4(1):10. doi: 10.1186/1756-8935-4-10. Epigenetics Chromatin. 2011. PMID: 21749726 Free PMC article.
-
Genomic imprinting of Dopa decarboxylase in heart and reciprocal allelic expression with neighboring Grb10.Mol Cell Biol. 2008 Jan;28(1):386-96. doi: 10.1128/MCB.00862-07. Epub 2007 Oct 29. Mol Cell Biol. 2008. PMID: 17967881 Free PMC article.
-
Genomic imprinting mechanisms in embryonic and extraembryonic mouse tissues.Heredity (Edinb). 2010 Jul;105(1):45-56. doi: 10.1038/hdy.2010.23. Epub 2010 Mar 17. Heredity (Edinb). 2010. PMID: 20234385 Free PMC article. Review.
-
Transcription factor ASCL2 is required for development of the glycogen trophoblast cell lineage.PLoS Genet. 2018 Aug 10;14(8):e1007587. doi: 10.1371/journal.pgen.1007587. eCollection 2018 Aug. PLoS Genet. 2018. PMID: 30096149 Free PMC article.
-
Postnatal survival of mice with maternal duplication of distal chromosome 7 induced by a Igf2/H19 imprinting control region lacking insulator function.PLoS Genet. 2010 Jan;6(1):e1000803. doi: 10.1371/journal.pgen.1000803. Epub 2010 Jan 8. PLoS Genet. 2010. PMID: 20062522 Free PMC article.
References
-
- McGrath J., Solter D. Completion of mouse embryogenesis requires both the maternal and paternal genomes. Cell. 1984;37:179–183. - PubMed
-
- Surani A., Barton S.C., Norris M.L. Development of reconstituted mouse eggs suggests imprinting of the genome during gametogenesis. Nature. 1984;308:548–550. - PubMed
-
- Morison I.M., Ramsay J.P., Spencer H.G. A census of mammalian imprinting. Trends Genet. 2005;21:457–465. - PubMed
-
- Barlow D.P. Gametic imprinting in mammals. Science. 1995;270:1610–1613. - PubMed
-
- Riordan D. Bioinformatic Analysis of Imprinted CpG Islands in Mus Musculus. Cambridge; 2003. M.Phil. thesis, Wellcome Trust Sanger Institute. http://www.sanger.ac.uk/Info/theses/
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
