

Dynamic molecular confinement in the plasma membrane by microdomains and the cytoskeleton meshwork
- PMID: 16858413
- PMCID: PMC1523176
- DOI: 10.1038/sj.emboj.7601214
Dynamic molecular confinement in the plasma membrane by microdomains and the cytoskeleton meshwork
Abstract
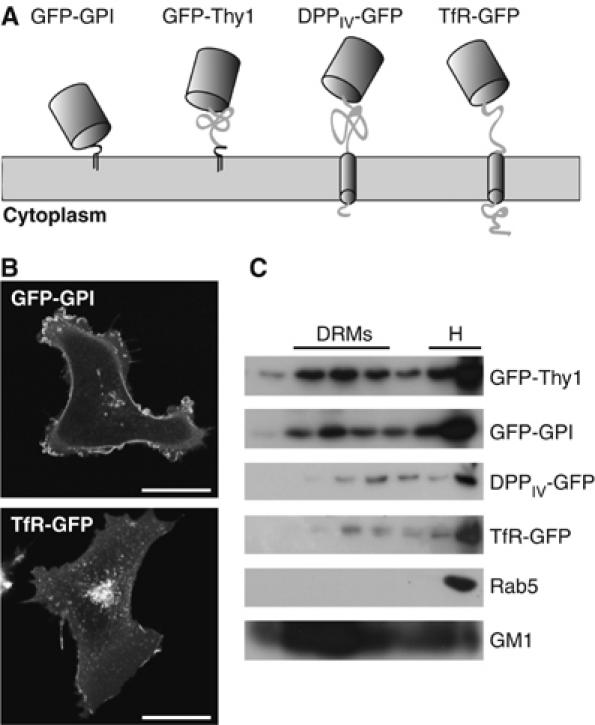
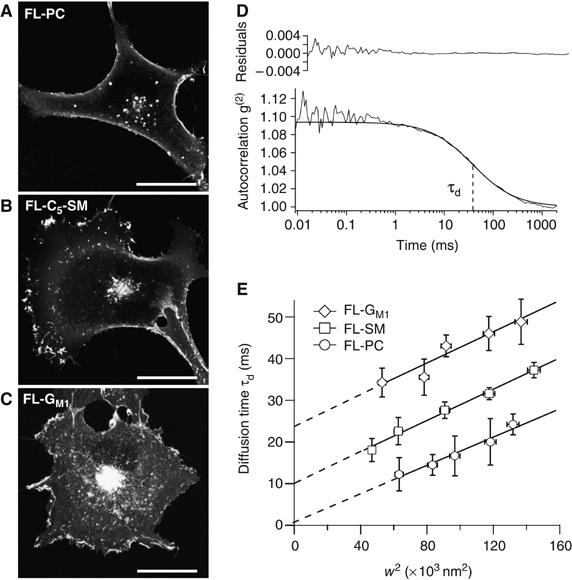
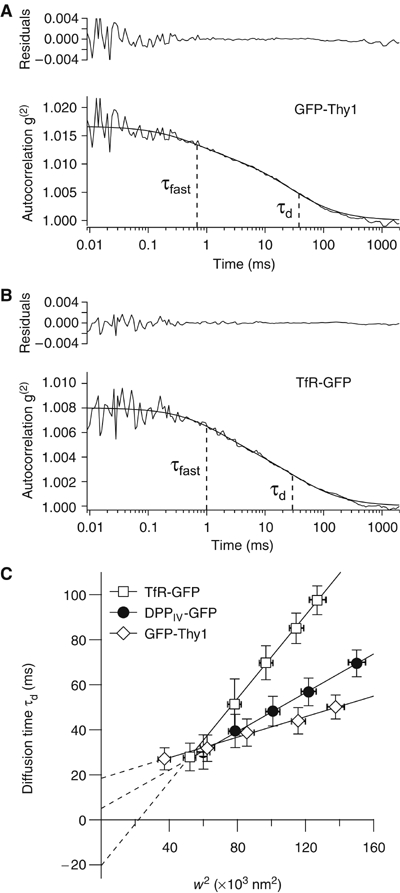
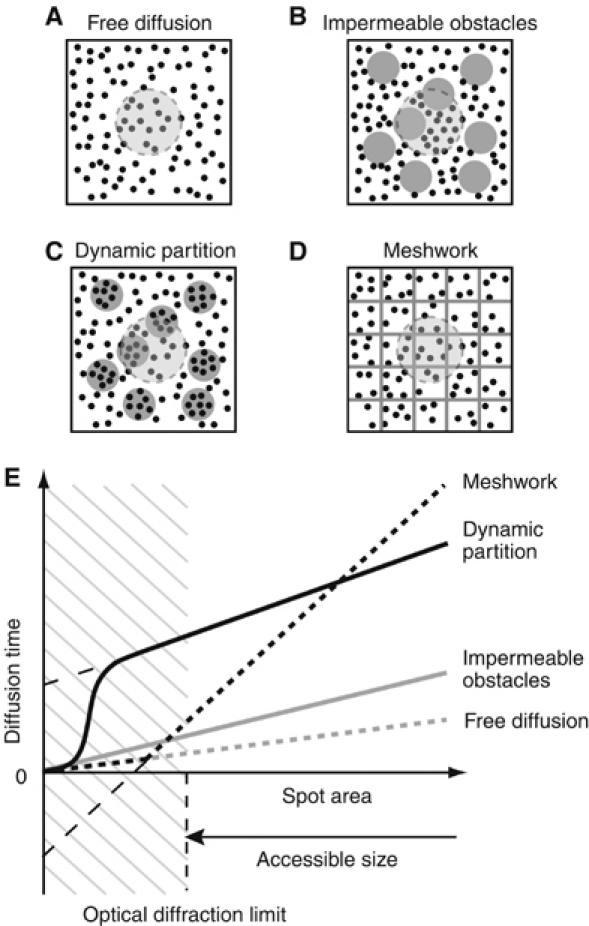
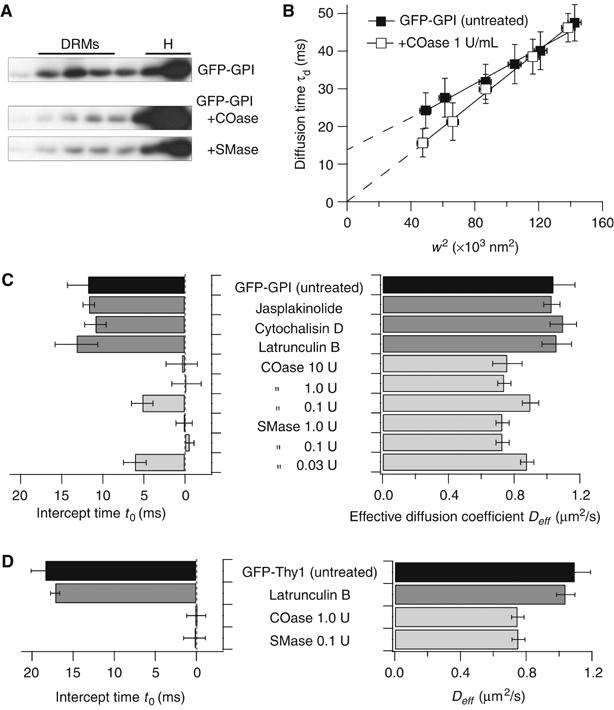
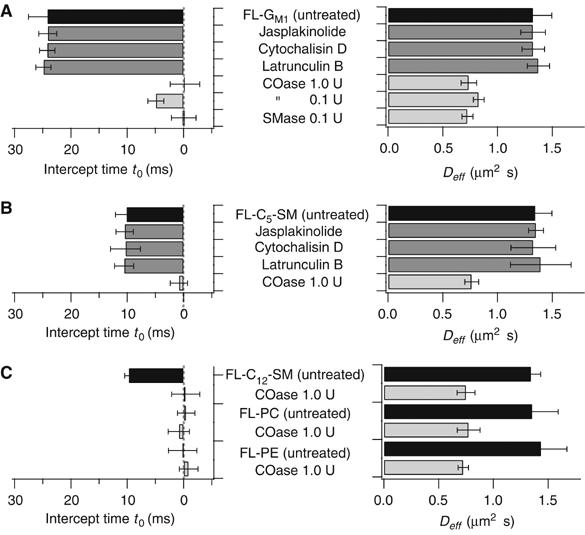
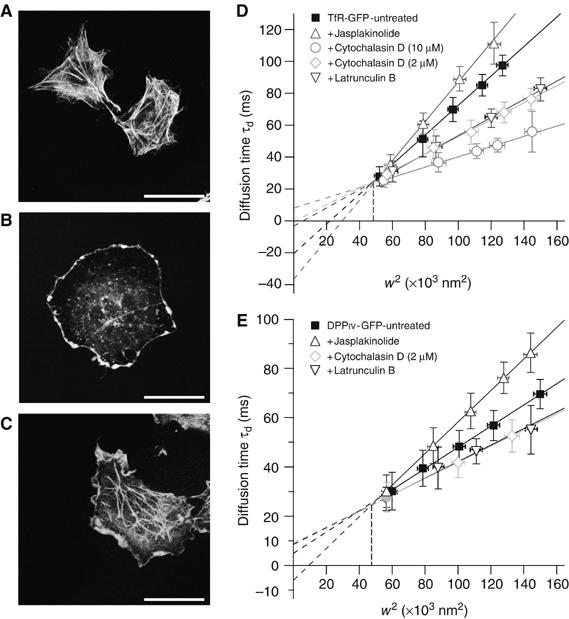
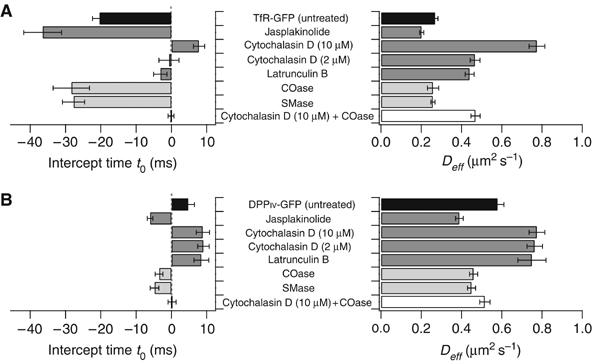
It is by now widely recognized that cell membranes show complex patterns of lateral organization. Two mechanisms involving either a lipid-dependent (microdomain model) or cytoskeleton-based (meshwork model) process are thought to be responsible for these plasma membrane organizations. In the present study, fluorescence correlation spectroscopy measurements on various spatial scales were performed in order to directly identify and characterize these two processes in live cells with a high temporal resolution, without any loss of spatial information. Putative raft markers were found to be dynamically compartmented within tens of milliseconds into small microdomains (Ø <120 nm) that are sensitive to the cholesterol and sphingomyelin levels, whereas actin-based cytoskeleton barriers are responsible for the confinement of the transferrin receptor protein. A free-like diffusion was observed when both the lipid-dependent and cytoskeleton-based organizations were disrupted, which suggests that these are two main compartmentalizing forces at work in the plasma membrane.
Figures
References
-
- Bligh EG, Dyer WJ (1959) A rapid method of total lipid extraction and purification. Can J Biochem Physiol 37: 911–917 - PubMed
-
- Brown DA, London E (1998) Functions of lipid rafts in biological membranes. Annu Rev Cell Dev Biol 14: 111–136 - PubMed
-
- Edidin M (2003) The state of lipid rafts: from model membranes to cells. Annu Rev Biophys Biomol Struct 32: 257–283 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
