

Immunity to recombinant plasmodium falciparum merozoite surface protein 1 (MSP1): protection in Aotus nancymai monkeys strongly correlates with anti-MSP1 antibody titer and in vitro parasite-inhibitory activity
- PMID: 16861644
- PMCID: PMC1539572
- DOI: 10.1128/IAI.01679-05
Immunity to recombinant plasmodium falciparum merozoite surface protein 1 (MSP1): protection in Aotus nancymai monkeys strongly correlates with anti-MSP1 antibody titer and in vitro parasite-inhibitory activity
Abstract
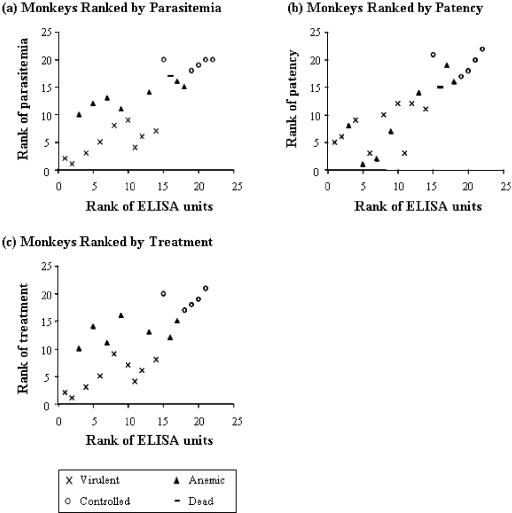
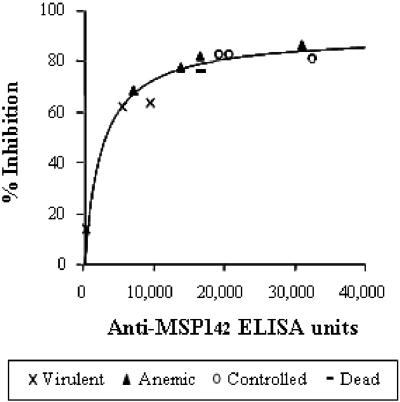
A number of malarial blood-stage candidate vaccines are currently being tested in human clinical trials, but our understanding of the relationship between clinical immunity and data obtained from in vitro assays remains inadequate. An in vitro assay which could reliably predict protective immunity in vivo would facilitate vaccine development. Merozoite surface protein1 (MSP1) is a leading blood-stage malaria vaccine candidate, and anti-MSP1 antibodies from individuals that are clinically immune to malaria inhibit the invasion of Plasmodium merozoites into erythrocytes in vitro. Using expression in Escherichia coli and subsequent refolding, we have produced two allelic forms of MSP1(42) (FVO and 3D7). Aotus nancymai monkeys were immunized with MSP1(42)-FVO, MSP1(42)-3D7, or a combination of FVO and 3D7 allelic forms, (MSP1(42)-C1) and were subsequently challenged with Plasmodium falciparum FVO parasites. Sera obtained prior to challenge were tested by standardized enzyme-linked immunosorbent assay (ELISA) to determine antibody titer, and immunoglobulin G (IgG) fractions were also obtained from the same sera; the IgG fractions were tested in an in vitro growth inhibition (GI) assay to evaluate biological activity of the antibodies. Regardless of the immunogen used, all monkeys that had >200,000 ELISA units against MSP1(42)-FVO antigen before challenge controlled their infections. By contrast, all monkeys whose purified IgGs gave <60% inhibition activity in an in vitro GI assay with P. falciparum FVO required treatment for high parasitemia after challenge. There is a strong correlation between ELISA units (Spearman rank correlation of greater than 0.75) or GI activity (Spearman rank correlation of greater than 0.70) and protective immunity judged by various parameters (e.g., cumulative parasitemia or day of patency). These data indicate that, in this monkey model, the ELISA and GI assay values can significantly predict protective immunity induced by a blood-stage vaccine, and they support the use of these assays as part of evaluation of human clinical trials of MSP1-based vaccines.
Figures

References
-
- Blackman, M. J., and A. A. Holder. 1992. Secondary processing of the Plasmodium falciparum merozoite surface protein-1 (MSP1) by a calcium-dependent membrane-bound serine protease: shedding of MSP133 as a noncovalently associated complex with other fragments of the MSP1. Mol. Biochem. Parasitol. 50:307-315. - PubMed
-
- Blackman, M. J., I. T. Ling, S. C. Nicholls, and A. A. Holder. 1991. Proteolytic processing of the Plasmodium falciparum merozoite surface protein-1 produces a membrane-bound fragment containing two epidermal growth factor-like domains. Mol. Biochem. Parasitol. 49:29-33. - PubMed
-
- Blackman, M. J., H. Whittle, and A. A. Holder. 1991. Processing of the Plasmodium falciparum major merozoite surface protein-1: identification of a 33-kilodalton secondary processing product which is shed prior to erythrocyte invasion. Mol. Biochem. Parasitol. 49:35-44. - PubMed
-
- Chang, S. P., S. E. Case, W. L. Gosnell, A. Hashimoto, K. J. Kramer, L. Q. Tam, C. Q. Hashiro, C. M. Nikaido, H. L. Gibson, C. T. Lee-Ng, P. J. Barr, B. T. Yokota, and G. S. Hut. 1996. A recombinant baculovirus 42-kilodalton C-terminal fragment of Plasmodium falciparum merozoite surface protein 1 protects Aotus monkeys against malaria. Infect. Immun. 64:253-261. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
