

Neural stem cells rescue nervous purkinje neurons by restoring molecular homeostasis of tissue plasminogen activator and downstream targets
- PMID: 16870729
- PMCID: PMC6674224
- DOI: 10.1523/JNEUROSCI.1624-06.2006
Neural stem cells rescue nervous purkinje neurons by restoring molecular homeostasis of tissue plasminogen activator and downstream targets
Abstract
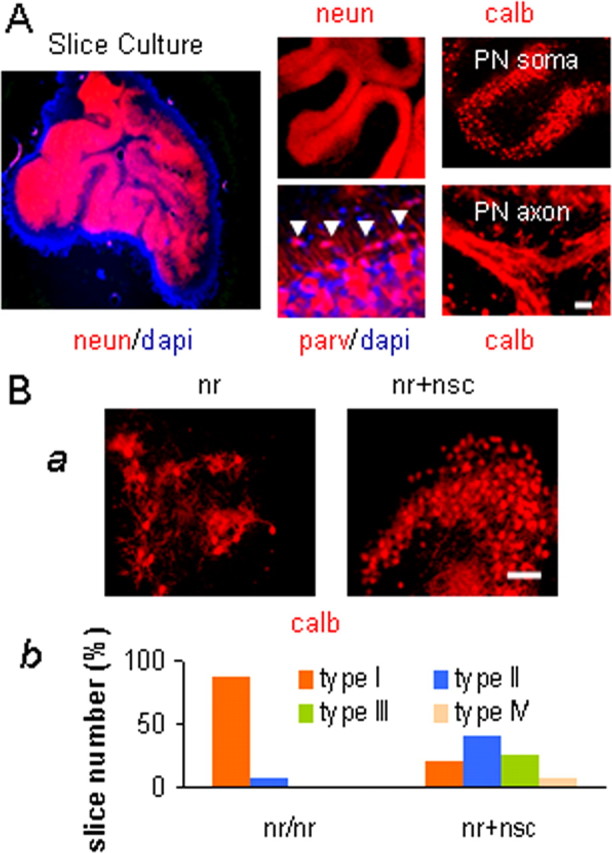
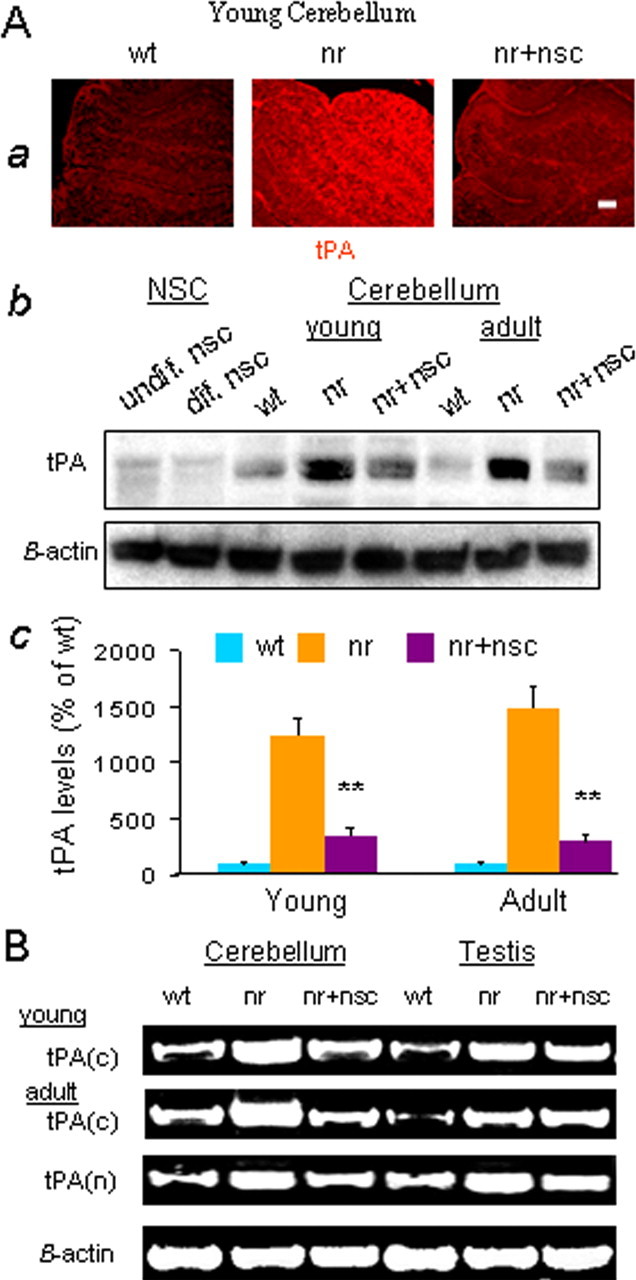
Neural stem cells (NSCs) offer special therapeutic prospects because they can be isolated from the CNS, expanded ex vivo, and re-implanted into diseased CNS where they not only migrate and differentiate according to cues from host tissue but also appear to be capable of affecting host cells. In nervous (nr) mutant mice Purkinje neuron (PN) mitochondria become abnormal by the second postnatal week, and a majority of PNs die in the fourth to fifth weeks. We previously identified in nr cerebellum a 10-fold increase in tissue plasminogen activator (tPA) as a key component of the mechanism causing nr PN death. Here we report that undifferentiated wild-type murine NSCs, when transplanted into the newborn nr cerebellar cortex, do not replace host PNs but contact imperiled PNs and support their mitochondrial function, dendritic growth, and synaptogenesis, subsequently leading to the rescue of host PNs and restoration of motor coordination. This protection of nr PNs also is verified by an in vitro organotypic slice model in which nr cerebellar slices are cocultured with NSCs. Most importantly, the integrated NSCs in young nr cerebellum rectify excessive tPA mRNA and protein to close to normal levels and protect the mitochondrial voltage-dependent anion channel and neurotrophins, downstream targets of the tPA/plasmin proteolytic system. This report demonstrates for the first time that NSCs can rescue imperiled host neurons by rectifying their gene expression, elevating somatic stem cell therapeutic potential beyond solely cell replacement strategy.
Figures





Similar articles
-
Purkinje neuron degeneration in nervous (nr) mutant mice is mediated by a metabolic pathway involving excess tissue plasminogen activator.Proc Natl Acad Sci U S A. 2006 May 16;103(20):7847-52. doi: 10.1073/pnas.0602440103. Epub 2006 May 8. Proc Natl Acad Sci U S A. 2006. PMID: 16682647 Free PMC article.
-
Tissue plasminogen activator regulates Purkinje neuron development and survival.Proc Natl Acad Sci U S A. 2013 Jun 25;110(26):E2410-9. doi: 10.1073/pnas.1305010110. Epub 2013 May 14. Proc Natl Acad Sci U S A. 2013. PMID: 23674688 Free PMC article.
-
Organotypic slices culture model for cerebellar ataxia: potential use to study Purkinje cell induction from neural stem cells.Brain Res Bull. 2011 Feb 1;84(2):169-73. doi: 10.1016/j.brainresbull.2010.12.001. Epub 2010 Dec 8. Brain Res Bull. 2011. PMID: 21145378
-
Neural stem cell biology may be well suited for improving brain tumor therapies.Cancer J. 2003 May-Jun;9(3):189-204. doi: 10.1097/00130404-200305000-00007. Cancer J. 2003. PMID: 12952304 Review.
-
Bcl-2 protection of axotomized Purkinje cells in organotypic culture is age dependent and not associated with an enhancement of axonal regeneration.Prog Brain Res. 2005;148:37-44. doi: 10.1016/S0079-6123(04)48004-2. Prog Brain Res. 2005. PMID: 15661179 Review. No abstract available.
Cited by
-
Experimental neurotransplantation treatment for hereditary cerebellar ataxias.Cerebellum Ataxias. 2016 Apr 4;3:7. doi: 10.1186/s40673-016-0045-3. eCollection 2016. Cerebellum Ataxias. 2016. PMID: 27047666 Free PMC article. Review.
-
Delayed transplantation of precursor cell-derived astrocytes provides multiple benefits in a rat model of Parkinsons.EMBO Mol Med. 2014 Apr;6(4):504-18. doi: 10.1002/emmm.201302878. Epub 2014 Jan 29. EMBO Mol Med. 2014. PMID: 24477866 Free PMC article.
-
Duchenne muscular dystrophy: current cell therapies.Ther Adv Neurol Disord. 2015 Jul;8(4):166-77. doi: 10.1177/1756285615586123. Ther Adv Neurol Disord. 2015. PMID: 26136844 Free PMC article. Review.
-
p38 MAPK-Mediated Bmi-1 down-regulation and defective proliferation in ATM-deficient neural stem cells can be restored by Akt activation.PLoS One. 2011 Jan 28;6(1):e16615. doi: 10.1371/journal.pone.0016615. PLoS One. 2011. PMID: 21305053 Free PMC article.
-
Neuroprotection: the emerging concept of restorative neural stem cell biology for the treatment of neurodegenerative diseases.Curr Neuropharmacol. 2011 Jun;9(2):313-7. doi: 10.2174/157015911795596603. Curr Neuropharmacol. 2011. PMID: 22131940 Free PMC article.
References
-
- Banerjee J, Ghosh S (2004). Interaction of mitochondrial voltage-dependent anion channel from rat brain with plasminogen protein leads to partial closure of the channel. Biochim Biophys Acta 1663:6–8. - PubMed
-
- Chen ZL, Strickland S (1997). Neuronal death in the hippocampus is promoted by plasmin-catalyzed degradation of laminin. Cell 91:917–925. - PubMed
-
- Christopherson KS, Ullian EM, Stokes CC, Mullowney CE, Hell JW, Agah A, Lawler J, Mosher DF, Bornstein P, Barres BA (2005). Thrombospondins are astrocyte-secreted proteins that promote CNS synaptogenesis. Cell 120:421–433. - PubMed
-
- De Jager PL, Harvey D, Polydorides AD, Zuo J, Heintz N (1998). A high-resolution genetic map of the nervous locus on mouse chromosome 8. Genomics 48:346–353. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases