

Robust integration of motion information in the fly visual system revealed by single cell photoablation
- PMID: 16870735
- PMCID: PMC6674221
- DOI: 10.1523/JNEUROSCI.1327-06.2006
Robust integration of motion information in the fly visual system revealed by single cell photoablation
Abstract
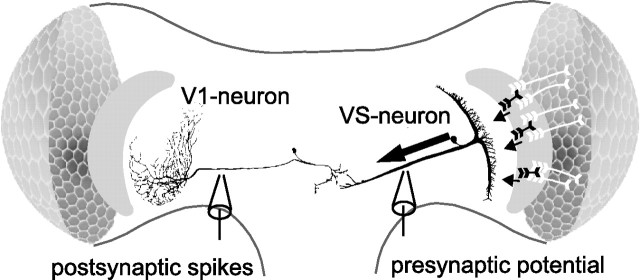
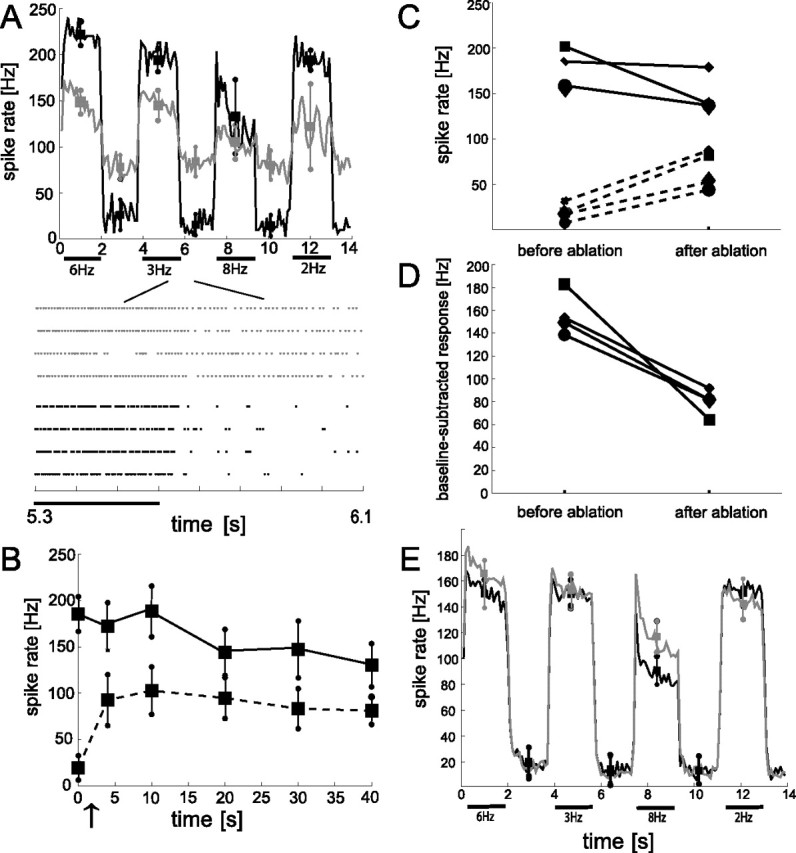
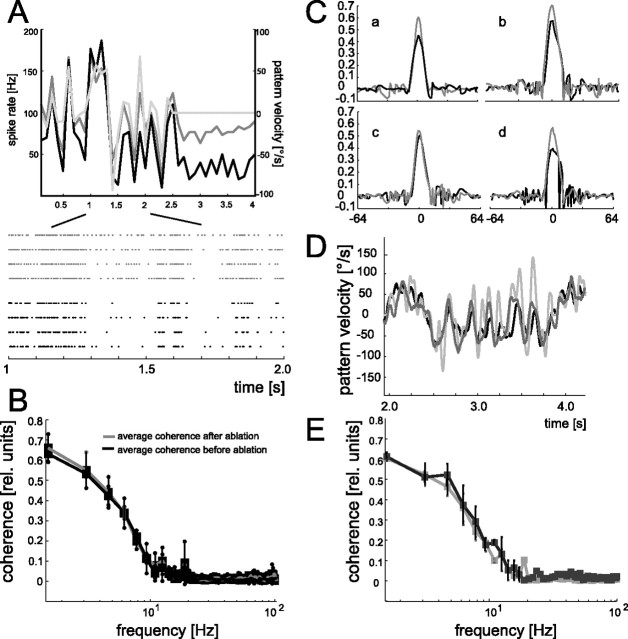
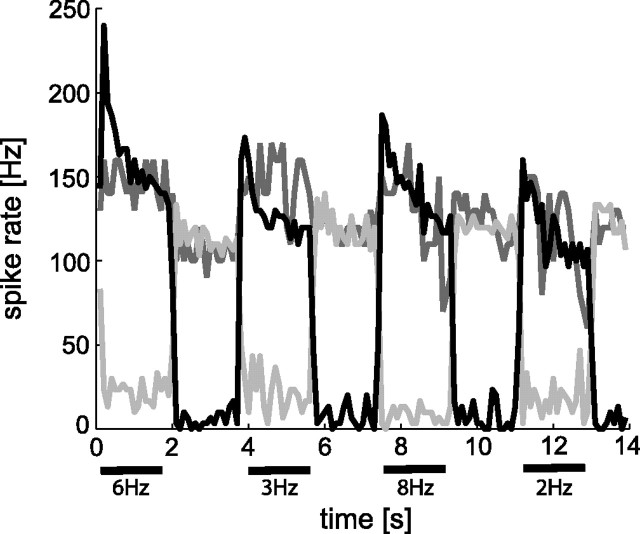
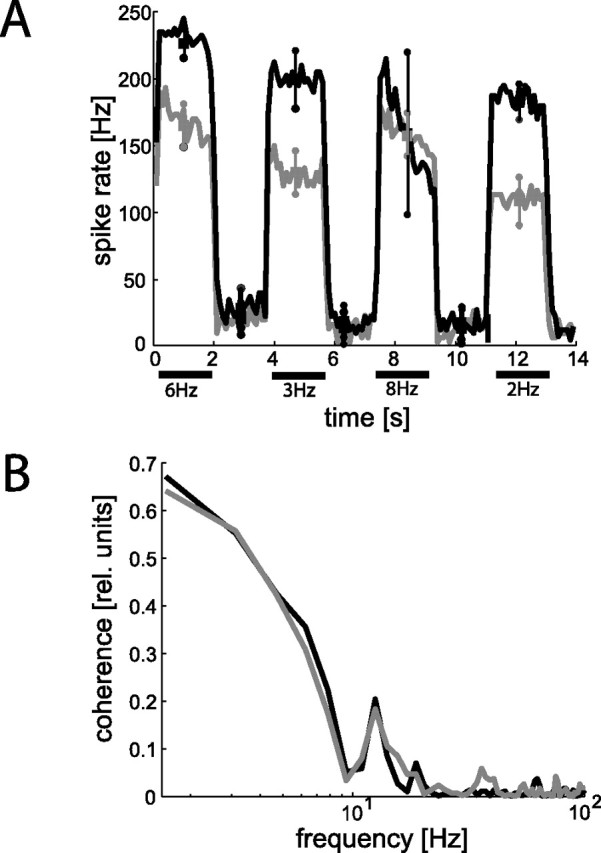
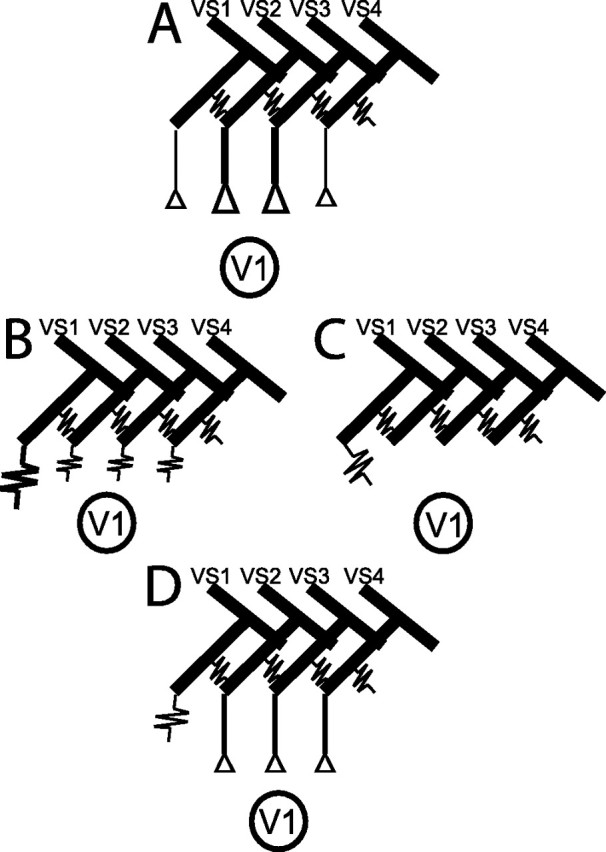
In the brain, sensory information needs often to be read out from the ensemble activity of presynaptic neurons. In the most basic case, this may be accomplished by an individual postsynaptic neuron. In the visual system of the blowfly, an identified motion-sensitive spiking neuron is known to be postsynaptic to an ensemble of graded-potential presynaptic input elements. Both the presynaptic and postsynaptic neurons were shown previously to be capable of representing the velocity of preferred-direction motion reliably and linearly over a large frequency range of velocity fluctuations. Accordingly, the synaptic transfer properties of the connecting excitatory synapses between individual input elements and the postsynaptic neuron were shown to be linear over a similar range of presynaptic membrane potential fluctuations. It was not known, however, how the postsynaptic neuron integrates and reads out the presynaptic ensemble activity. We were able to compare the response properties of the integrating cell before and after eliminating individual presynaptic elements by a laser ablation technique. For most of the input elements, we found that their elimination strongly affected the activity of the postsynaptic neuron but did not degrade its performance to encode motion with constant and time-varying velocity. Our results suggest that the integration of individual synaptic inputs within the neural circuit operates with some redundancy. This feature might help the postsynaptic neuron to encode in a highly robust way the direction and the velocity of self-motion of the animal.
Figures
Similar articles
-
Synaptic transfer of dynamic motion information between identified neurons in the visual system of the blowfly.Neuroscience. 2003;119(4):1103-12. doi: 10.1016/s0306-4522(03)00204-5. Neuroscience. 2003. PMID: 12831867
-
Transfer of visual motion information via graded synapses operates linearly in the natural activity range.J Neurosci. 2001 Sep 1;21(17):6957-66. doi: 10.1523/JNEUROSCI.21-17-06957.2001. J Neurosci. 2001. PMID: 11517283 Free PMC article.
-
Precise timing in fly motion vision is mediated by fast components of combined graded and spike signals.Neuroscience. 2009 May 19;160(3):639-50. doi: 10.1016/j.neuroscience.2009.02.045. Epub 2009 Mar 3. Neuroscience. 2009. PMID: 19264111
-
Neural encoding of behaviourally relevant visual-motion information in the fly.Trends Neurosci. 2002 Feb;25(2):96-102. doi: 10.1016/s0166-2236(02)02063-5. Trends Neurosci. 2002. PMID: 11814562 Review.
-
Common circuit design in fly and mammalian motion vision.Nat Neurosci. 2015 Aug;18(8):1067-76. doi: 10.1038/nn.4050. Epub 2015 Jun 29. Nat Neurosci. 2015. PMID: 26120965 Review.
Cited by
-
Adaptation of velocity encoding in synaptically coupled neurons in the fly visual system.J Neurosci. 2008 Sep 10;28(37):9183-93. doi: 10.1523/JNEUROSCI.1936-08.2008. J Neurosci. 2008. PMID: 18784299 Free PMC article.
-
Electrical coupling of lobula plate tangential cells to a heterolateral motion-sensitive neuron in the fly.J Neurosci. 2008 Dec 31;28(53):14435-42. doi: 10.1523/JNEUROSCI.3603-08.2008. J Neurosci. 2008. PMID: 19118177 Free PMC article.
-
Spatial vision in insects is facilitated by shaping the dynamics of visual input through behavioral action.Front Neural Circuits. 2012 Dec 20;6:108. doi: 10.3389/fncir.2012.00108. eCollection 2012. Front Neural Circuits. 2012. PMID: 23269913 Free PMC article.
-
Reciprocal inhibitory connections within a neural network for rotational optic-flow processing.Front Neurosci. 2007 Oct 15;1(1):111-21. doi: 10.3389/neuro.01.1.1.008.2007. eCollection 2007 Nov. Front Neurosci. 2007. PMID: 18982122 Free PMC article.
-
Neural basis of stimulus-angle-dependent motor control of wind-elicited walking behavior in the cricket Gryllus bimaculatus.PLoS One. 2013 Nov 14;8(11):e80184. doi: 10.1371/journal.pone.0080184. eCollection 2013. PLoS One. 2013. PMID: 24244644 Free PMC article.
References
-
- Borst A, Haag J (2002). Neural networks in the cockpit of the fly. J Comp Physiol A Neuroethol Sens Neural Behav Physiol 188:419–437. - PubMed
-
- Borst A, Theunissen FE (1999). Information theory and neural coding. Nat Neurosci 2:947–957. - PubMed
-
- Brenner N, Strong SP, Koberle R, Bialek W, de Ruyter van Steveninck RR (2000). Synergy in a neural code. Neural Comput 12:1531–1552. - PubMed
-
- de Ruyter van Steveninck RR, Laughlin SB (1996). The rate of information transfer at graded-potential synapses. Nature 379:642–645.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources