

Ubl4b, an X-derived retrogene, is specifically expressed in post-meiotic germ cells in mammals
- PMID: 16872915
- PMCID: PMC1647293
- DOI: 10.1016/j.modgep.2006.06.002
Ubl4b, an X-derived retrogene, is specifically expressed in post-meiotic germ cells in mammals
Abstract
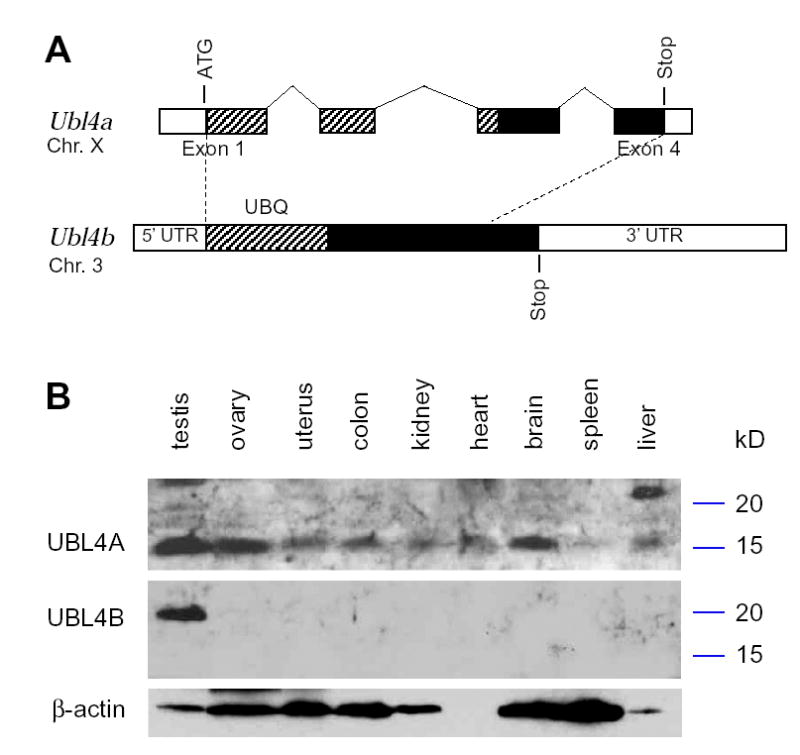
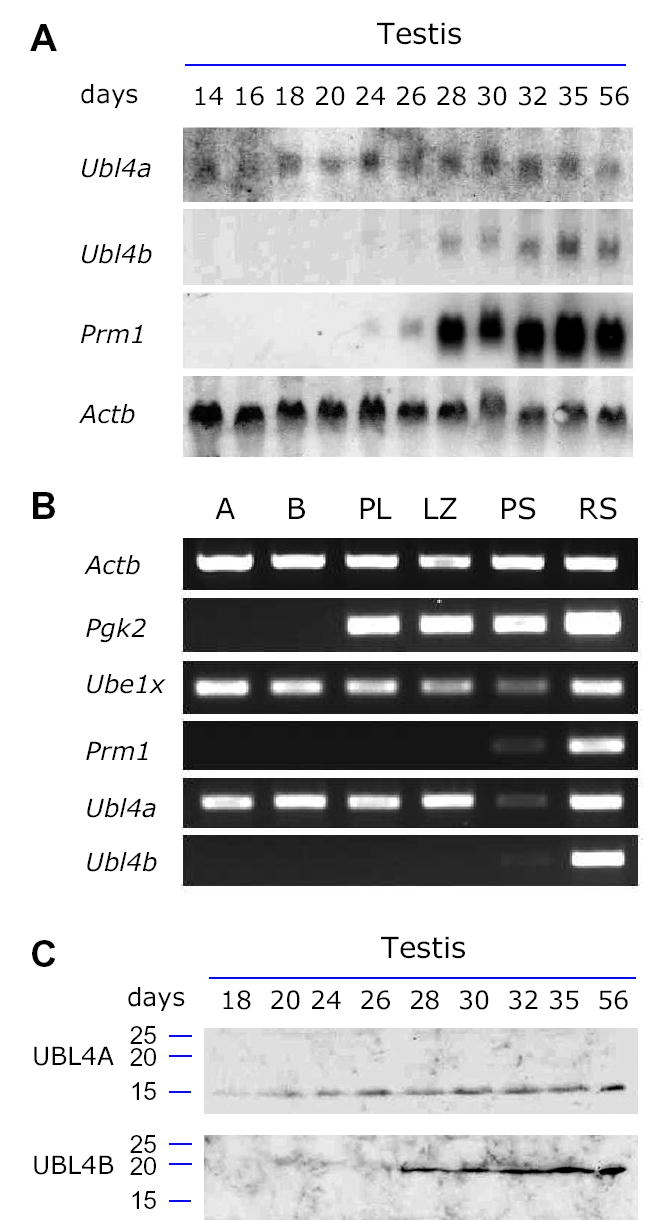
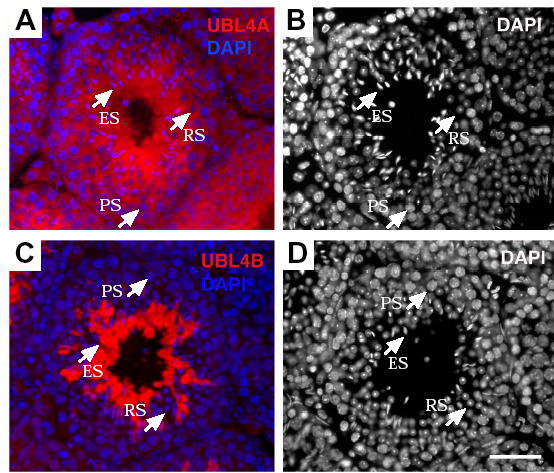
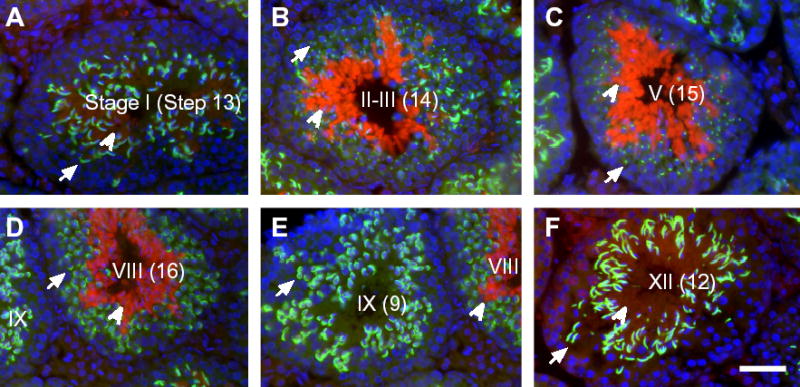
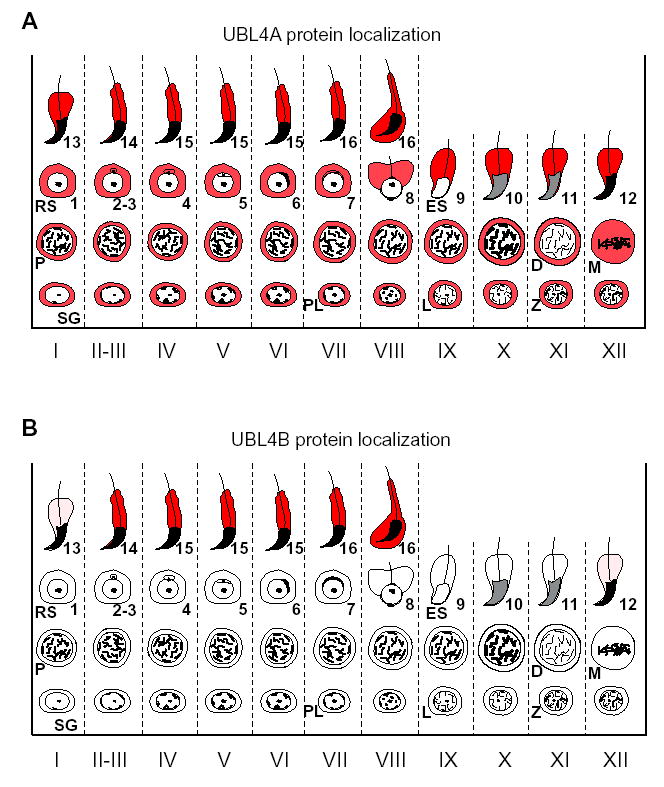
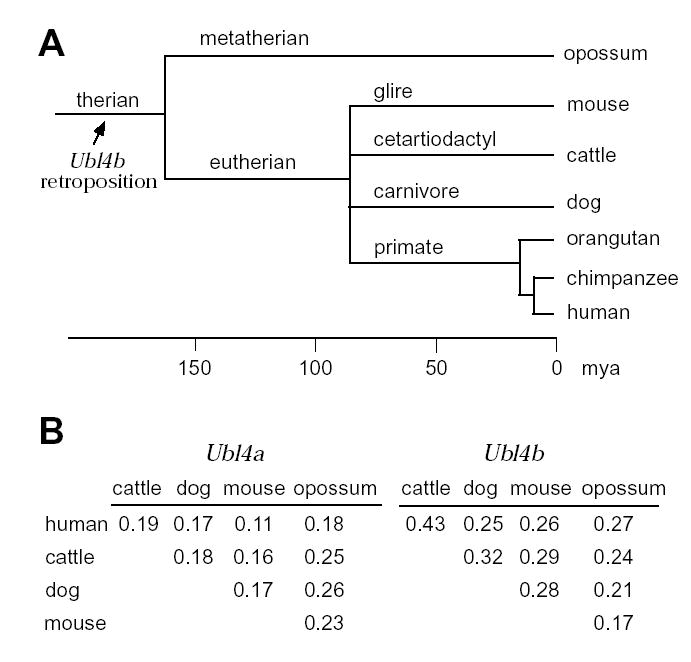
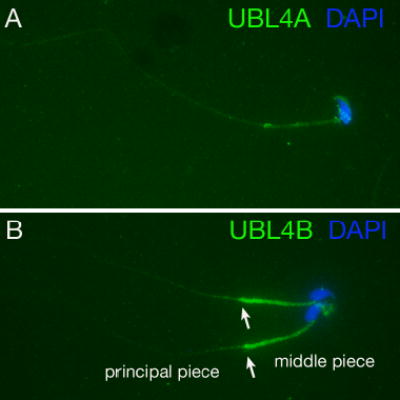
Post-translational modification by ubiquitin and ubiquitin-related proteins plays critical roles in protein degradation and in regulation of essential cellular processes. In mammals, transcription grinds to a halt during late spermiogenesis due to compaction of the spermatid genome, which creates a special need for robust post-transcriptional regulation. Here, we report the finding of a novel mouse ubiquitin-like protein, UBL4B. Ubl4b is a testis-specific autosomal gene. Ubl4b lacks introns and evidently arose from an X-linked intron-bearing housekeeping gene, Ubl4a, by retroposition during mammalian evolution. While Ubl4a is expressed throughout spermatogenesis, Ubl4b is restricted to post-meiotic germ cells. Ubl4a is highly conserved, but Ubl4b has undergone rapid evolution and may have evolved new functions. Our data suggest that evolution of Ubl4b is not due to meiotic sex chromosome inactivation (MSCI). Alternatively, origination of Ubl4b was due to MSCI, but Ubl4b eventually evolved to be restricted to post-meiotic germ cells.
Figures
References
-
- Bebington C, Doherty FJ, Fleming SD. The possible biological and reproductive functions of ubiquitin. Hum Reprod Update. 2001;7:102–111. - PubMed
-
- Bradley J, Baltus A, Skaletsky H, Royce-Tolland M, Dewar K, Page DC. An X-to-autosome retrogene is required for spermatogenesis in mice. Nat Genet. 2004;36:872–876. - PubMed
-
- Dahl HH, Brown RM, Hutchison WM, Maragos C, Brown GK. A testis-specific form of the human pyruvate dehydrogenase E1 alpha subunit is coded for by an intronless gene on chromosome 4. Genomics. 1990;8:225–232. - PubMed
-
- Dass B, McMahon KW, Jenkins NA, Gilbert DJ, Copeland NG, MacDonald CC. The gene for a variant form of the polyadenylation protein CstF-64 is on chromosome 19 and is expressed in pachytene spermatocytes in mice. J Biol Chem. 2001;276:8044–8050. - PubMed
-
- Elliott DJ, Venables JP, Newton CS, Lawson D, Boyle S, Eperon IC, Cooke HJ. An evolutionarily conserved germ cell-specific hnRNP is encoded by a retrotransposed gene. Hum Mol Genet. 2000;9:2117–2124. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
