

Amino acid residues in the N-terminal region of the PA subunit of influenza A virus RNA polymerase play a critical role in protein stability, endonuclease activity, cap binding, and virion RNA promoter binding
- PMID: 16873236
- PMCID: PMC1563815
- DOI: 10.1128/JVI.00600-06
Amino acid residues in the N-terminal region of the PA subunit of influenza A virus RNA polymerase play a critical role in protein stability, endonuclease activity, cap binding, and virion RNA promoter binding
Abstract
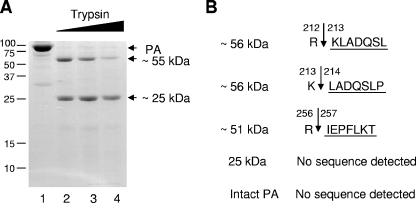
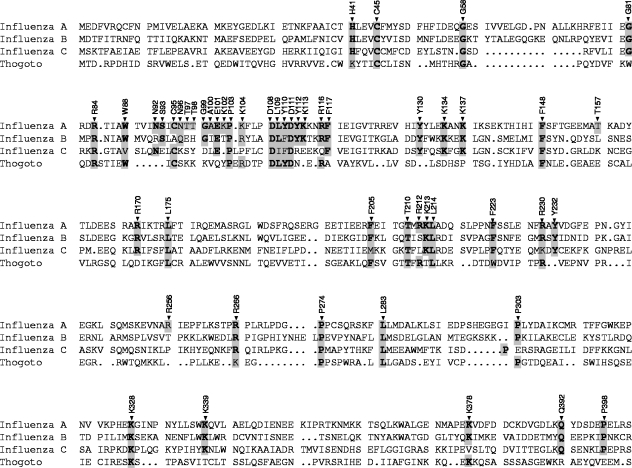
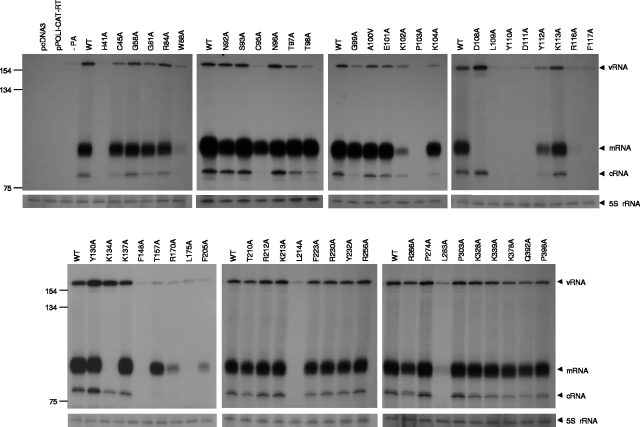
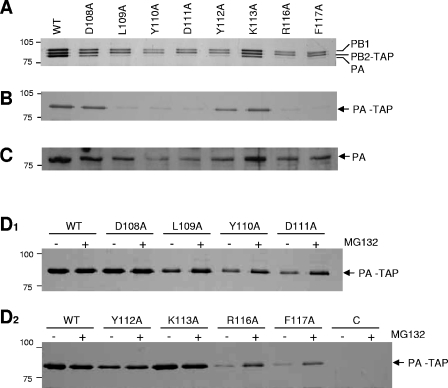
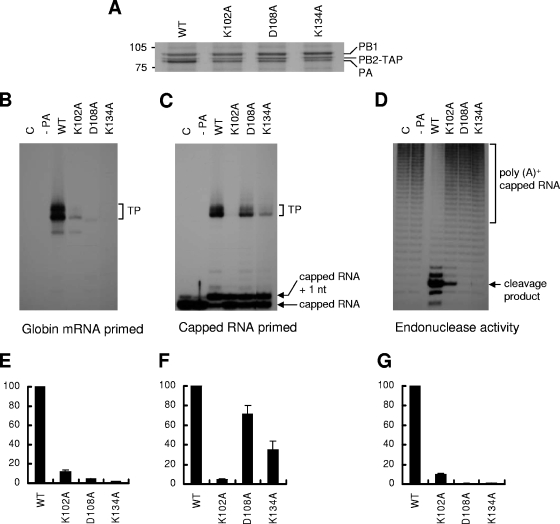
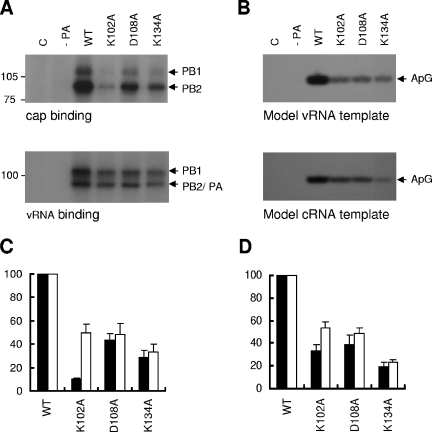
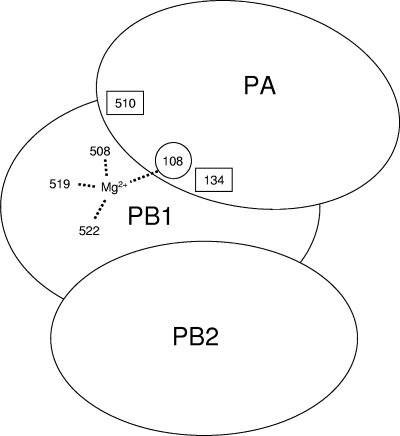
The RNA-dependent RNA polymerase of influenza virus is a heterotrimer formed by the PB1, PB2, and PA subunits. Although PA is known to be required for polymerase activity, its precise role is still unclear. Here, we investigated the function of the N-terminal region of PA. Protease digestion of purified recombinant influenza virus A/PR/8/34 PA initially suggested that its N-terminal region is folded into a 25-kDa domain. We then systematically introduced point mutations into evolutionarily conserved amino acids in the N-terminal region of influenza virus A/WSN/33. Most alanine-scanning mutations between residues L109 and F117 caused PA degradation, mediated by a proteasome-ubiquitin pathway, and as a consequence interfered with polymerase activity. Three further PA mutations, K102A, D108A, and K134A, were investigated in detail. Mutation K102A caused a general decrease both in transcription and replication in vivo, whereas mutations D108A and K134A selectively inhibited transcription. Both the D108A and K134A mutations completely inhibited endonuclease activity in vitro, explaining their selective defect in transcription. K102A, on the other hand, resulted in a significant decrease in both cap binding and viral RNA promoter-binding activity and consequently inhibited both transcription and replication. These results suggest that the N-terminal region of PA is involved in multiple functions of the polymerase, including protein stability, endonuclease activity, cap binding, and promoter binding.
Figures
Similar articles
-
PA and PA-X: two key proteins from segment 3 of the influenza viruses.Front Cell Infect Microbiol. 2025 Mar 14;15:1560250. doi: 10.3389/fcimb.2025.1560250. eCollection 2025. Front Cell Infect Microbiol. 2025. PMID: 40160474 Free PMC article. Review.
-
A single amino acid mutation in the PA subunit of the influenza virus RNA polymerase inhibits endonucleolytic cleavage of capped RNAs.J Virol. 2002 Sep;76(18):8989-9001. doi: 10.1128/jvi.76.18.8989-9001.2002. J Virol. 2002. PMID: 12186883 Free PMC article.
-
The cap-snatching endonuclease of influenza virus polymerase resides in the PA subunit.Nature. 2009 Apr 16;458(7240):914-8. doi: 10.1038/nature07745. Epub 2009 Feb 4. Nature. 2009. PMID: 19194459
-
Crystal structure of an avian influenza polymerase PA(N) reveals an endonuclease active site.Nature. 2009 Apr 16;458(7240):909-13. doi: 10.1038/nature07720. Epub 2009 Feb 4. Nature. 2009. PMID: 19194458
-
[Structural and Biochemical Analyses on the RNA-dependent RNA Polymerase of Influenza Virus for Development of Novel Anti-influenza Agents].Yakugaku Zasshi. 2017;137(2):205-214. doi: 10.1248/yakushi.16-00195. Yakugaku Zasshi. 2017. PMID: 28154333 Review. Japanese.
Cited by
-
Recent Advances in the Phytochemistry of Bryophytes: Distribution, Structures and Biological Activity of Bibenzyl and Bisbibenzyl Compounds.Plants (Basel). 2023 Dec 15;12(24):4173. doi: 10.3390/plants12244173. Plants (Basel). 2023. PMID: 38140499 Free PMC article. Review.
-
A Eurasian avian-like H1N1 swine influenza reassortant virus became pathogenic and highly transmissible due to mutations in its PA gene.Proc Natl Acad Sci U S A. 2022 Aug 23;119(34):e2203919119. doi: 10.1073/pnas.2203919119. Epub 2022 Aug 15. Proc Natl Acad Sci U S A. 2022. PMID: 35969783 Free PMC article.
-
Artificial hybrids of influenza A virus RNA polymerase reveal PA subunit modulates its thermal sensitivity.PLoS One. 2010 Dec 7;5(12):e15140. doi: 10.1371/journal.pone.0015140. PLoS One. 2010. PMID: 21151876 Free PMC article.
-
Selective Degradation of Host RNA Polymerase II Transcripts by Influenza A Virus PA-X Host Shutoff Protein.PLoS Pathog. 2016 Feb 5;12(2):e1005427. doi: 10.1371/journal.ppat.1005427. eCollection 2016 Feb. PLoS Pathog. 2016. PMID: 26849127 Free PMC article.
-
PA and PA-X: two key proteins from segment 3 of the influenza viruses.Front Cell Infect Microbiol. 2025 Mar 14;15:1560250. doi: 10.3389/fcimb.2025.1560250. eCollection 2025. Front Cell Infect Microbiol. 2025. PMID: 40160474 Free PMC article. Review.
References
-
- Blok, V., C. Cianci, K. W. Tibbles, S. C. Inglis, M. Krystal, and P. Digard. 1996. Inhibition of the influenza virus RNA-dependent RNA polymerase by antisera directed against the carboxy-terminal region of the PB2 subunit. J. Gen. Virol. 77:1025-1033. - PubMed
-
- Brownlee, G. G., E. Fodor, D. C. Pritlove, K. G. Gould, and J. J. Dalluge. 1995. Solid phase synthesis of 5′-diphosphorylated oligoribonucleotides and their conversion to capped m7Gppp-oligoribonucleotides for use as primers for influenza A virus RNA polymerase in vitro. Nucleic Acids Res. 23:2641-2647. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
