

Mechanisms underlying beta2-adrenoceptor-mediated nitric oxide generation by human umbilical vein endothelial cells
- PMID: 16873402
- PMCID: PMC1890348
- DOI: 10.1113/jphysiol.2006.115998
Mechanisms underlying beta2-adrenoceptor-mediated nitric oxide generation by human umbilical vein endothelial cells
Abstract
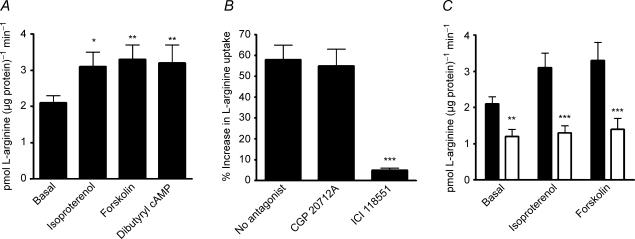
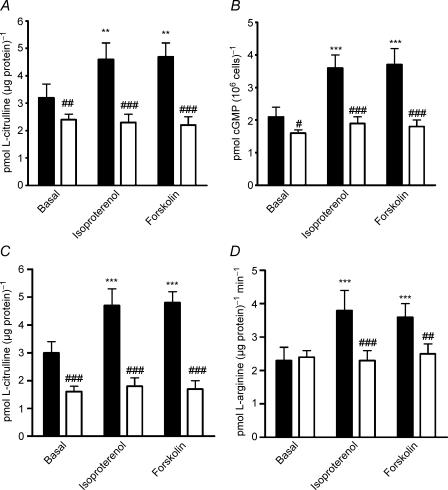
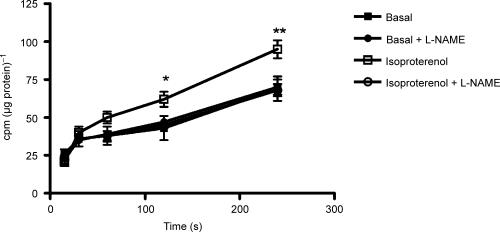
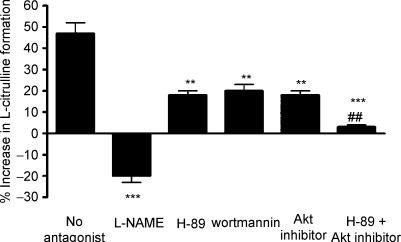
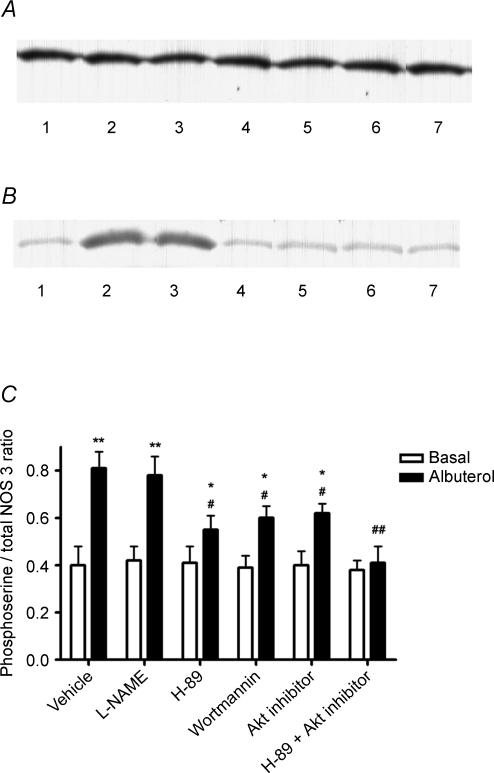
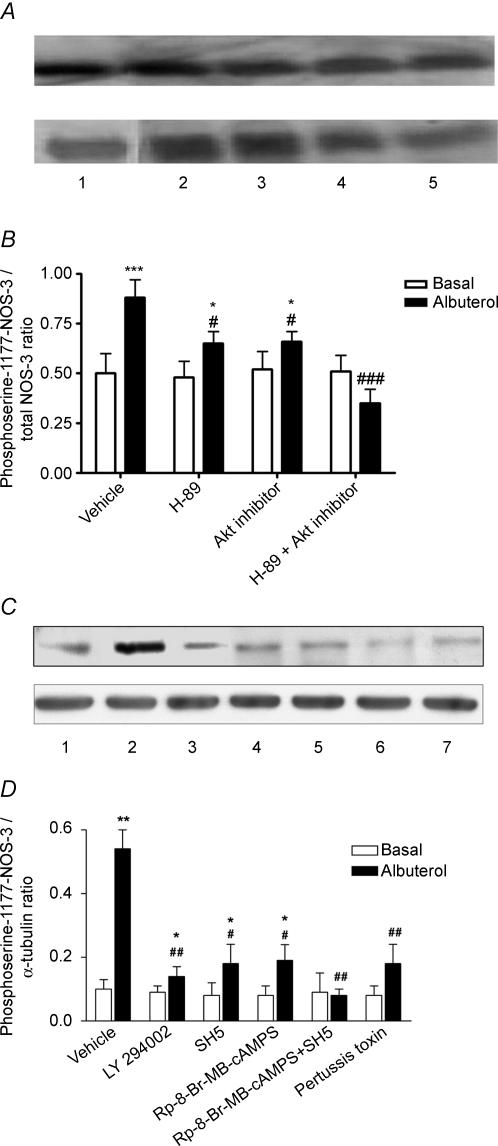
Endothelial beta(2)-adrenoceptor (beta(2)AR) stimulation increases nitric oxide (NO) generation, but the underlying cellular mechanisms are unclear. We examined the role of l-arginine transport and of phosphorylation of NO synthase 3 (NOS-3) in beta(2)AR-mediated NO biosynthesis by human umbilical vein endothelial cells (HUVEC). To this end, we assessed l-arginine uptake, NOS activity (from l-arginine to l-citrulline conversion), membrane potential (using [(3)H]tetraphenylphosphonium), as well as serine phosphorylation of NOS-3 (by Western blotting and mass spectrometry), in HUVEC treated with betaAR agonists or cyclic AMP-elevating agents. beta(2)AR stimulation increased l-arginine transport, as did cyclic AMP elevation with either forskolin or dibutyryl cyclic AMP, and this increase was inhibitable by N-ethylmaleimide. Blockade of l-arginine uptake by l-lysine inhibited NOS activity and, conversely, blockade of NOS using N(omega)-nitro-l-arginine methyl ester (l-NAME) inhibited l-arginine transport. beta(2)AR stimulation also caused a membrane hyperpolarization inhibitable by l-NAME, suggesting that the increase in l-arginine uptake occurred in response to NO-mediated hyperpolarization. beta(2)AR activation also increased NOS activity and phosphorylation of NOS-3 on serine-1177, and these increases were attenuated by inhibition of protein kinase A (PKA), phosphatidylinositol 3-kinase (PI3K) or Akt, and abolished by coinhibition of PKA and Akt. These findings suggest that beta(2)AR-mediated NOS-3 activation in HUVEC is mediated through phosphorylation of NOS-3 on serine-1177 through both the PKA and the PI3K/Akt systems, and is sustained by an increase in l-arginine uptake resulting from NO-mediated membrane hyperpolarization.
Figures

Similar articles
-
Activation of nitric oxide synthase by beta 2-adrenoceptors in human umbilical vein endothelium in vitro.Br J Pharmacol. 1999 Apr;126(8):1872-80. doi: 10.1038/sj.bjp.0702512. Br J Pharmacol. 1999. PMID: 10372832 Free PMC article.
-
Nitric oxide-dependent beta2-adrenergic dilatation of rat aorta is mediated through activation of both protein kinase A and Akt.Br J Pharmacol. 2004 Oct;143(3):397-403. doi: 10.1038/sj.bjp.0705933. Epub 2004 Sep 6. Br J Pharmacol. 2004. PMID: 15351777 Free PMC article.
-
Early activation of the p42/p44MAPK pathway mediates adenosine-induced nitric oxide production in human endothelial cells: a novel calcium-insensitive mechanism.FASEB J. 2002 Oct;16(12):1584-94. doi: 10.1096/fj.01-0125com. FASEB J. 2002. PMID: 12374781
-
A role for insulin on L-arginine transport in fetal endothelial dysfunction in hyperglycaemia.Curr Vasc Pharmacol. 2009 Oct;7(4):467-74. doi: 10.2174/157016109789043919. Curr Vasc Pharmacol. 2009. PMID: 19485892 Review.
-
Arginine metabolism and the synthesis of nitric oxide in the nervous system.Prog Neurobiol. 2001 Jul;64(4):365-91. doi: 10.1016/s0301-0082(00)00056-3. Prog Neurobiol. 2001. PMID: 11275358 Review.
Cited by
-
Role of endothelial cells in antihyperalgesia induced by a triptan and β-blocker.Neuroscience. 2013 Mar 1;232:83-9. doi: 10.1016/j.neuroscience.2012.12.020. Epub 2012 Dec 20. Neuroscience. 2013. PMID: 23262231 Free PMC article.
-
Role of reactive oxygen species in hyperadrenergic hypertension: biochemical, physiological, and pharmacological evidence from targeted ablation of the chromogranin a (Chga) gene.Circ Cardiovasc Genet. 2010 Oct;3(5):414-25. doi: 10.1161/CIRCGENETICS.109.924050. Epub 2010 Aug 20. Circ Cardiovasc Genet. 2010. PMID: 20729505 Free PMC article.
-
Colforsin-induced vasodilation in chronic hypoxic pulmonary hypertension in rats.J Anesth. 2010 Jun;24(3):432-40. doi: 10.1007/s00540-010-0912-7. Epub 2010 Mar 19. J Anesth. 2010. PMID: 20300779
-
Anti-inflammatory effects of β2 adrenergic receptor agonists in experimental acute lung injury.FASEB J. 2012 May;26(5):2137-44. doi: 10.1096/fj.11-201640. Epub 2012 Feb 7. FASEB J. 2012. PMID: 22318967 Free PMC article.
-
β Adrenergic Receptor Kinase C-Terminal Peptide Gene-Therapy Improves β2-Adrenergic Receptor-Dependent Neoangiogenesis after Hindlimb Ischemia.J Pharmacol Exp Ther. 2016 Feb;356(2):503-13. doi: 10.1124/jpet.115.228411. Epub 2015 Nov 24. J Pharmacol Exp Ther. 2016. PMID: 26604244 Free PMC article.
References
-
- Arcangeli A, Olivotto M. Plasma membrane potential of murine erythroleukemia cells: approach to measurement and evidence for cell-density dependence. J Cell Physiol. 1986;127:17–27. - PubMed
-
- Bae SW, Kim HS, Cha YN, Park YS, Jo SA, Jo I. Rapid increase in endothelial nitric oxide production by bradykinin is mediated by protein kinase A signaling pathway. Biochem Biophys Res Commun. 2003;306:981–987. - PubMed
-
- Boo YC, Sorescu G, Boyd N, Shiojima I, Walsh K, Du J, et al. Shear stress stimulates phosphorylation of endothelial nitric-oxide synthase at Ser1179 by Akt-independent mechanisms: role of protein kinase A. J Biol Chem. 2002;277:3388–3396. - PubMed
-
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–254. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials