

Local and diffuse mechanisms of primary afferent depolarization and presynaptic inhibition in the rat spinal cord
- PMID: 16873417
- PMCID: PMC1995647
- DOI: 10.1113/jphysiol.2006.110577
Local and diffuse mechanisms of primary afferent depolarization and presynaptic inhibition in the rat spinal cord
Abstract
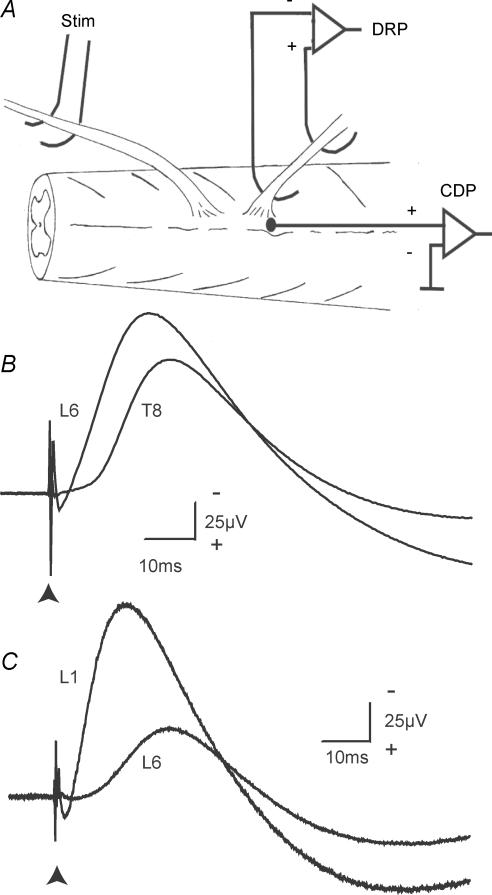
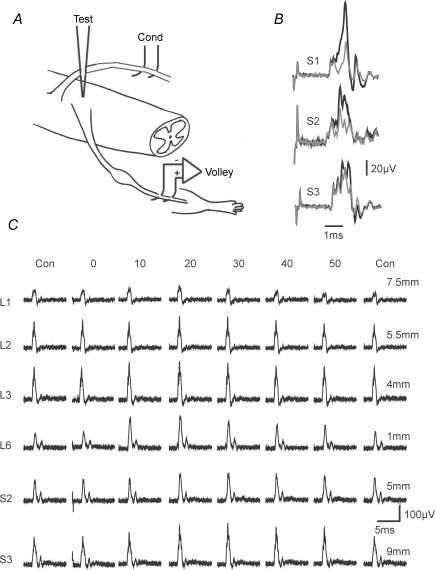
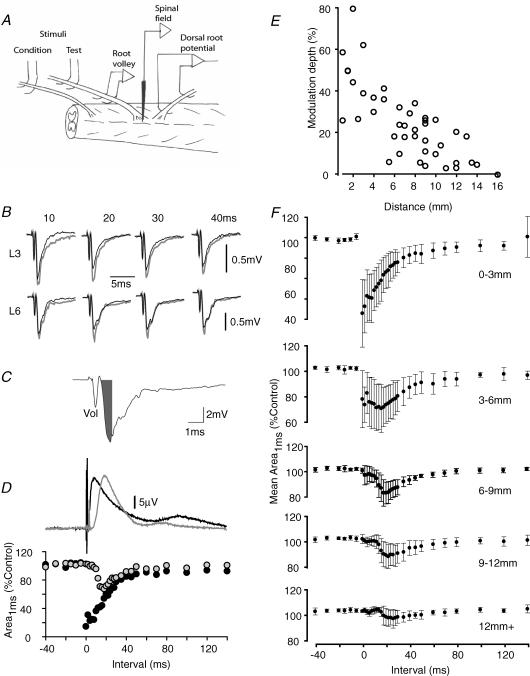
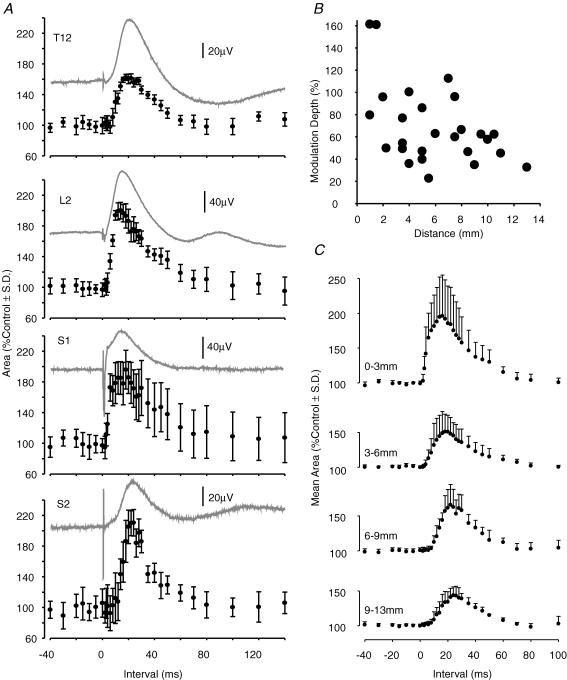
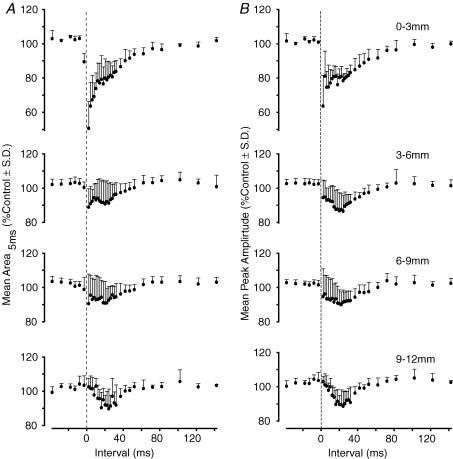
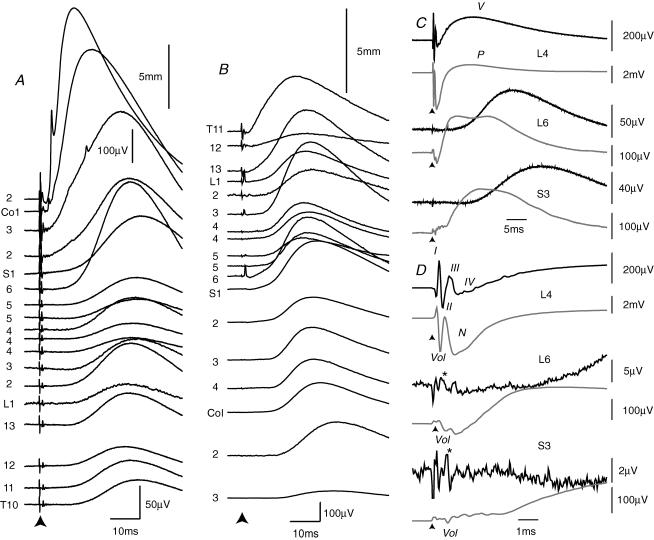
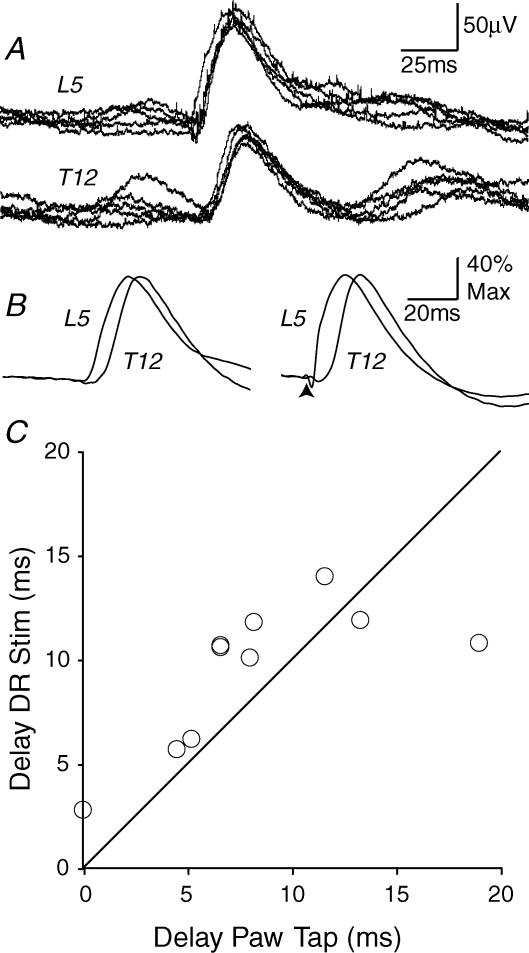
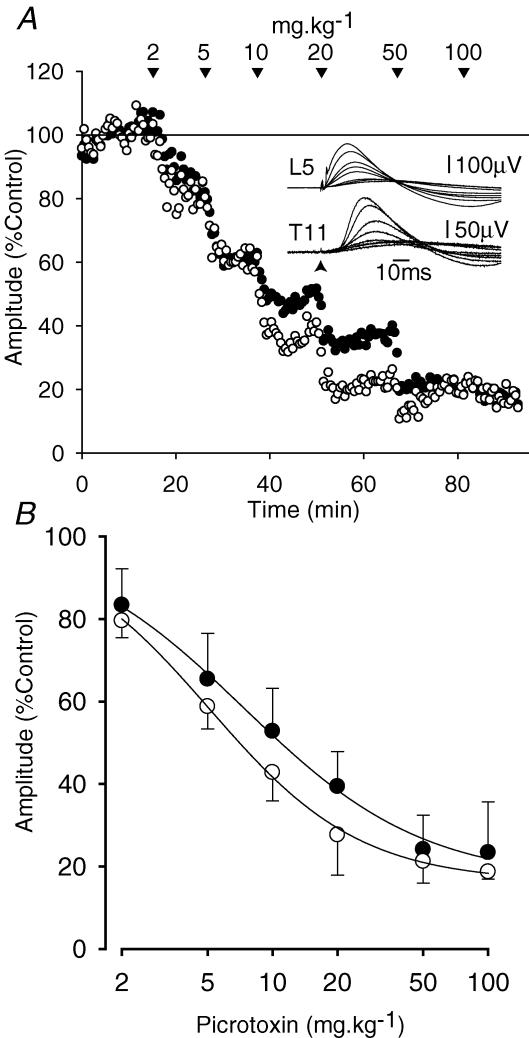
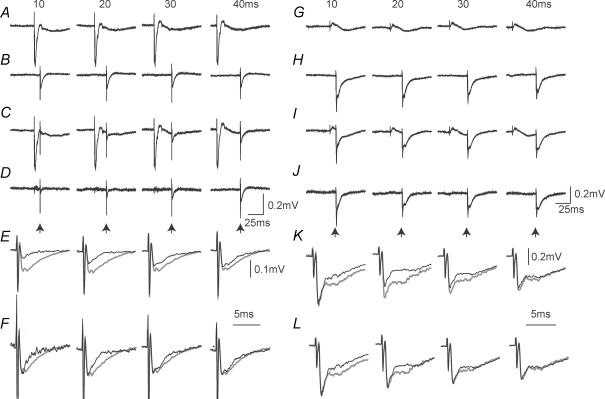
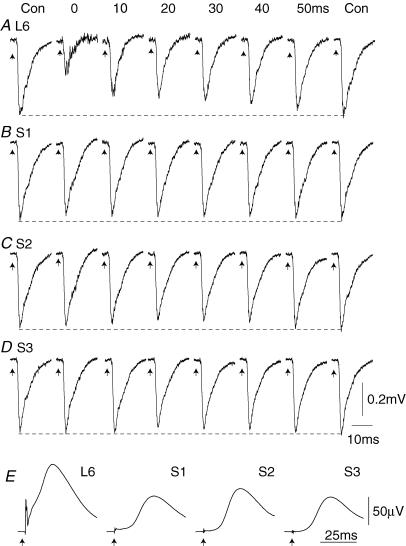
Two types of dorsal root potential (DRP) were found in the spinal cord of urethane-anaesthetized rats. Local DRPs with short latency-to-onset were evoked on roots close to the point of entry of an afferent volley. Diffuse DRPs with a longer latency-to-onset were seen on more distant roots up to 17 segments from the volley entry zone. The switch to long latency-to-onset occurred abruptly as a function of distance along the cord and could not be explained by conduction delays within the dorsal columns. Long-latency DRPs were also present and superimposed on the short-latency DRPs on nearby roots. Both local and diffuse DRPs were evoked by light mechanical stimuli: von Frey hair thresholds were <or= 1 gram force Changes in excitability of the terminals of sural nerve afferents were used to confirm that both local and diffuse DRPs were associated with primary afferent depolarization (PAD). These effects were potent: the area of the antidromic volley evoked in the sural nerve by intraspinal microstimulation in the L4/5 spinal segment was increased by 109 +/- 50% (mean +/- s.d.; n = 5) by nearby conditioning stimuli, and by 52 +/- 12% (n = 6) with stimuli applied 9-13 mm (5-8 segments) away. The time course of the changes in terminal excitability closely matched those of the DRPs. Reduction of the field potentials evoked in the dorsal horn by stimulation of dorsal roots was also shown to accompany both local and diffuse DRPs. The area of the monosynaptically evoked field potential was reduced by 48 +/- 19% (n = 7) with nearby conditioning stimulation and 16 +/- 9% (n = 10) with stimulation 9-12 mm distant. Evidence is presented that this inhibition includes a presynaptic component. Similar effects were seen with field potentials evoked by sural nerve stimulation. It is concluded that diffuse DRPs are mediated through propriospinal networks which may contribute to the gating of sensory information flow during natural behaviour as they respond to weak mechanical stimuli and provoke presynaptic inhibition.
Figures
Similar articles
-
Five sources of a dorsal root potential: their interactions and origins in the superficial dorsal horn.J Neurophysiol. 1997 Aug;78(2):860-71. doi: 10.1152/jn.1997.78.2.860. J Neurophysiol. 1997. PMID: 9307119
-
Muscle afferent excitability testing in spinal root-intact rats: dissociating peripheral afferent and efferent volleys generated by intraspinal microstimulation.J Neurophysiol. 2017 Feb 1;117(2):796-807. doi: 10.1152/jn.00874.2016. Epub 2016 Dec 14. J Neurophysiol. 2017. PMID: 27974451 Free PMC article.
-
Interneurones mediating presynaptic inhibition of group II muscle afferents in the cat spinal cord.J Physiol. 1995 Mar 1;483 ( Pt 2)(Pt 2):461-71. doi: 10.1113/jphysiol.1995.sp020597. J Physiol. 1995. PMID: 7650613 Free PMC article.
-
Antidromic discharges of dorsal root afferents in the neonatal rat.J Physiol Paris. 1999 Sep-Oct;93(4):359-67. doi: 10.1016/s0928-4257(00)80063-7. J Physiol Paris. 1999. PMID: 10574124 Review.
-
Origins of antidromic activity in sensory afferent fibers and neurogenic inflammation.Semin Immunopathol. 2018 May;40(3):237-247. doi: 10.1007/s00281-017-0669-2. Epub 2018 Feb 8. Semin Immunopathol. 2018. PMID: 29423889 Free PMC article. Review.
Cited by
-
Elucidating afferent-driven presynaptic inhibition of primary afferent input to spinal laminae I and X.Front Cell Neurosci. 2023 Jan 11;16:1029799. doi: 10.3389/fncel.2022.1029799. eCollection 2022. Front Cell Neurosci. 2023. PMID: 36713779 Free PMC article.
-
Lateral inhibition during nociceptive processing.Pain. 2017 Jun;158(6):1046-1052. doi: 10.1097/j.pain.0000000000000876. Pain. 2017. PMID: 28195854 Free PMC article.
-
Monoaminergic modulation of spinal viscero-sympathetic function in the neonatal mouse thoracic spinal cord.PLoS One. 2012;7(11):e47213. doi: 10.1371/journal.pone.0047213. Epub 2012 Nov 5. PLoS One. 2012. PMID: 23144807 Free PMC article.
-
Post-activation depression from primary afferent depolarization (PAD) produces extensor H-reflex suppression following flexor afferent conditioning.J Physiol. 2023 May;601(10):1925-1956. doi: 10.1113/JP283706. Epub 2023 Apr 9. J Physiol. 2023. PMID: 36928599 Free PMC article.
-
Depressing effect of electroacupuncture on the spinal non-painful sensory input of the rat.Exp Brain Res. 2014 Sep;232(9):2721-9. doi: 10.1007/s00221-014-3965-2. Epub 2014 Apr 26. Exp Brain Res. 2014. PMID: 24770863
References
-
- Bayev KV, Kostyuk PG. Primary afferent depolarization evoked by the activity of spinal scratching generator. Neuroscience. 1981;6:205–215. - PubMed
-
- Beloozerova I, Rossignol S. Antidromic discharges in dorsal roots of decerebrate cats – I. Studies at rest and during fictive locomotion. Brain Res. 1999;846:87–105. - PubMed
-
- Beloozerova IN, Rossignol S. Antidromic discharges in dorsal roots of decerebrate cats. II: studies during treadmill locomotion. Brain Res. 2004;996:227–236. - PubMed
-
- Besson JM, Rivot JP. Heterosegmental, heterosensory and cortical inhibitory effects on dorsal interneurones in the cat's spinal cord. Electroencephalogr Clin Neurophysiol. 1972;33:195–206. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources