

The potential regulation of L1 mobility by RNA interference
- PMID: 16877813
- PMCID: PMC1559915
- DOI: 10.1155/JBB/2006/32713
The potential regulation of L1 mobility by RNA interference
Abstract
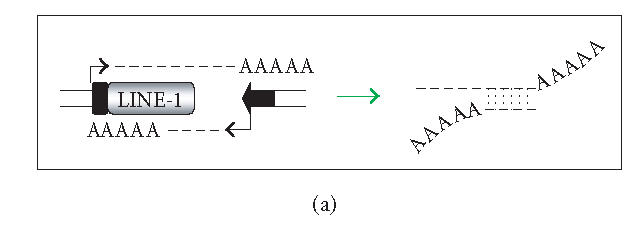
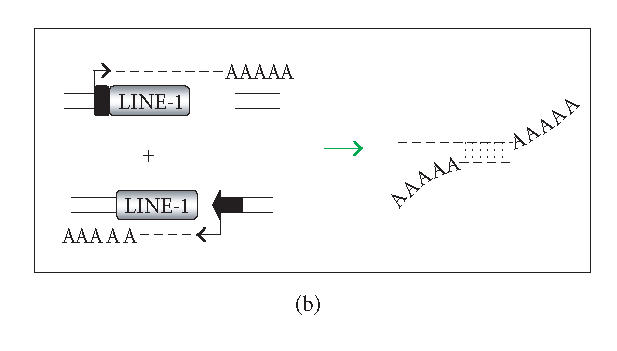
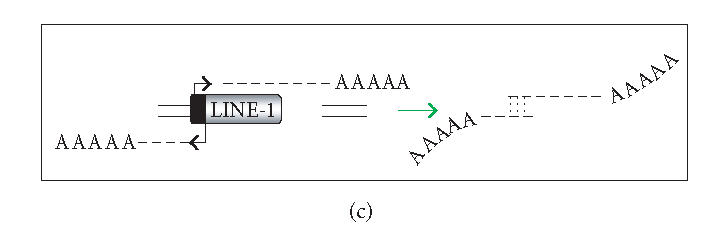
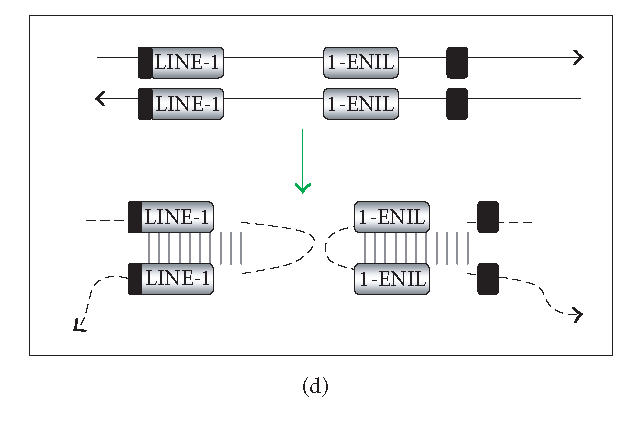
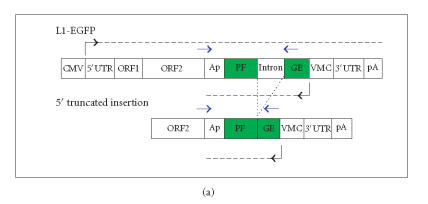
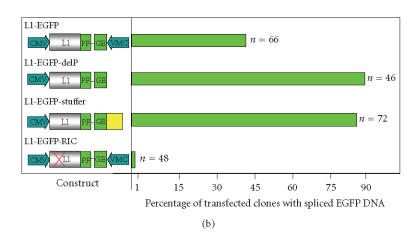
The hypothesis that RNA interference constrains L1 mobility seems inherently reasonable: L1 mobility can be dangerous and L1 RNA, the presumed target of RNAi, serves as a critical retrotransposition intermediate. Despite its plausibility, proof for this hypothesis has been difficult to obtain. Studies attempting to link the L1 retrotransposition frequency to alterations in RNAi activity have been hampered by the long times required to measure retrotransposition frequency, the pleiotropic and toxic effects of altering RNAi over similar time periods, and the possibility that other cellular machinery may contribute to the regulation of L1s. Another problem is that the commonly used L1 reporter cassette may serve as a substrate for RNAi. Here we review the L1-RNAi hypothesis and describe a genetic assay with a modified reporter cassette that detects approximately 4 times more L1 insertions than the conventional retrotransposition assay.
Figures
Similar articles
-
L1 retrotransposition is suppressed by endogenously encoded small interfering RNAs in human cultured cells.Nat Struct Mol Biol. 2006 Sep;13(9):763-71. doi: 10.1038/nsmb1141. Epub 2006 Aug 27. Nat Struct Mol Biol. 2006. PMID: 16936727
-
A potential role for RNA interference in controlling the activity of the human LINE-1 retrotransposon.Nucleic Acids Res. 2005 Feb 8;33(3):846-56. doi: 10.1093/nar/gki223. Print 2005. Nucleic Acids Res. 2005. PMID: 15701756 Free PMC article.
-
Human L1 retrotransposition: insights and peculiarities learned from a cultured cell retrotransposition assay.Genetica. 1999;107(1-3):39-51. Genetica. 1999. PMID: 10952196 Review.
-
Gamma radiation increases endonuclease-dependent L1 retrotransposition in a cultured cell assay.Nucleic Acids Res. 2006 Feb 28;34(4):1196-204. doi: 10.1093/nar/gkj522. Print 2006. Nucleic Acids Res. 2006. PMID: 16507671 Free PMC article.
-
Biology of mammalian L1 retrotransposons.Annu Rev Genet. 2001;35:501-38. doi: 10.1146/annurev.genet.35.102401.091032. Annu Rev Genet. 2001. PMID: 11700292 Review.
Cited by
-
ZCCHC3 is a stress granule zinc knuckle protein that strongly suppresses LINE-1 retrotransposition.PLoS Genet. 2023 Jul 5;19(7):e1010795. doi: 10.1371/journal.pgen.1010795. eCollection 2023 Jul. PLoS Genet. 2023. PMID: 37405998 Free PMC article.
-
Restricted and non-essential redundancy of RNAi and piRNA pathways in mouse oocytes.PLoS Genet. 2019 Dec 20;15(12):e1008261. doi: 10.1371/journal.pgen.1008261. eCollection 2019 Dec. PLoS Genet. 2019. PMID: 31860668 Free PMC article.
-
Transposable elements: genome innovation, chromosome diversity, and centromere conflict.Chromosome Res. 2018 Mar;26(1-2):5-23. doi: 10.1007/s10577-017-9569-5. Epub 2018 Jan 13. Chromosome Res. 2018. PMID: 29332159 Free PMC article. Review.
-
Functional protein pathway activation mapping of the progression of normal skin to squamous cell carcinoma.Cancer Prev Res (Phila). 2012 Mar;5(3):403-13. doi: 10.1158/1940-6207.CAPR-11-0427. Cancer Prev Res (Phila). 2012. PMID: 22389437 Free PMC article.
References
-
- Tomari Y, Zamore PD. Perspective: machines for RNAi. Genes & Development. 2005;19(5):517–529. - PubMed
-
- Carmell MA, Hannon GJ. RNase III enzymes and the initiation of gene silencing. Nature Structural & Molecular Biology. 2004;11(3):214–218. - PubMed
-
- Plasterk RHA. RNA silencing: the genome's immune system. Science. 2002;296(5571):1263–1265. - PubMed
-
- Bagasra O, Prilliman KR. RNA interference: the molecular immune system. Journal of Molecular Histology. 2004;35(6):545–553. - PubMed
-
- Sijen T, Plasterk RHA. Transposon silencing in the Caenorhabditis elegans germ line by natural RNAi. Nature. 2003;426(6964):310–314. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
