

Ensemble molecular dynamics yields submillisecond kinetics and intermediates of membrane fusion
- PMID: 16880392
- PMCID: PMC1567673
- DOI: 10.1073/pnas.0601597103
Ensemble molecular dynamics yields submillisecond kinetics and intermediates of membrane fusion
Abstract
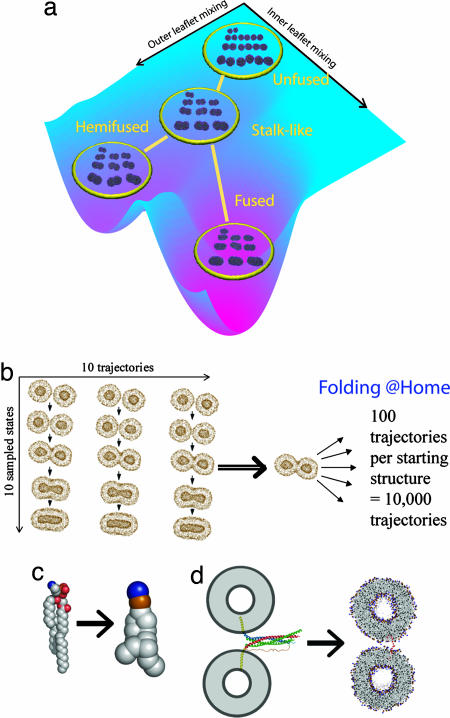
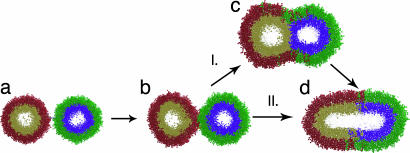
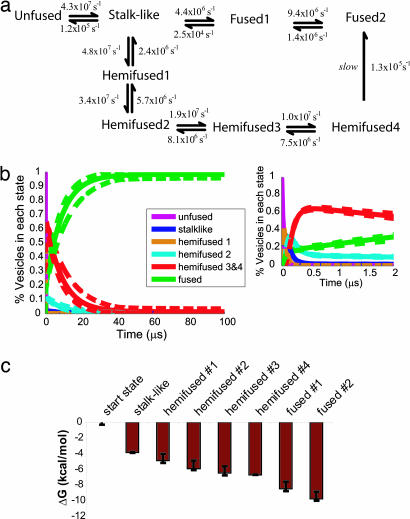
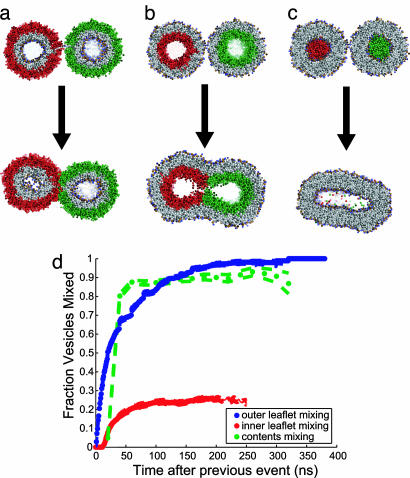
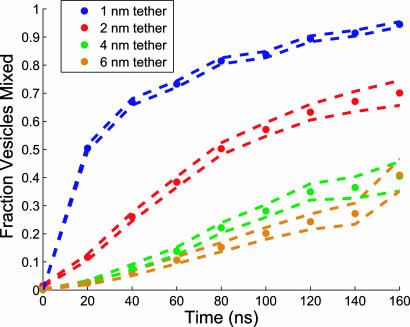
Lipid membrane fusion is critical to cellular transport and signaling processes such as constitutive secretion, neurotransmitter release, and infection by enveloped viruses. Here, we introduce a powerful computational methodology for simulating membrane fusion from a starting configuration designed to approximate activated prefusion assemblies from neuronal and viral fusion, producing results on a time scale and degree of mechanistic detail not previously possible to our knowledge. We use an approach to the long time scale simulation of fusion by constructing a Markovian state model with large-scale distributed computing, yielding an understanding of fusion mechanisms on time scales previously impossible to simulate to our knowledge. Our simulation data suggest a branched pathway for fusion, in which a common stalk-like intermediate can either rapidly form a fusion pore or remain in a metastable hemifused state that slowly forms fully fused vesicles. This branched reaction pathway provides a mechanistic explanation both for the biphasic fusion kinetics and the stable hemifused intermediates previously observed experimentally. Our distributed computing and Markovian state model approaches provide sufficient sampling to detect rare transitions, a systematic process for analyzing reaction pathways, and the ability to develop quantitative approximations of reaction kinetics for fusion.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
