

Biglycan binds to alpha- and gamma-sarcoglycan and regulates their expression during development
- PMID: 16883602
- PMCID: PMC2929672
- DOI: 10.1002/jcp.20740
Biglycan binds to alpha- and gamma-sarcoglycan and regulates their expression during development
Abstract
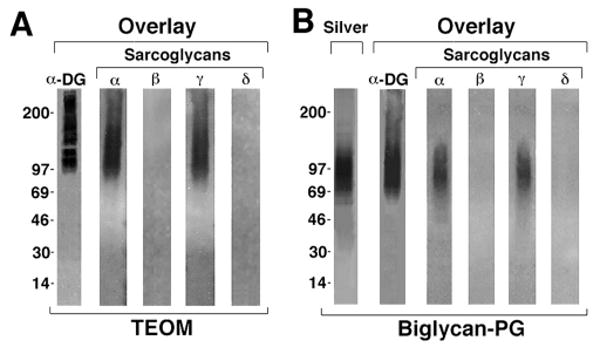
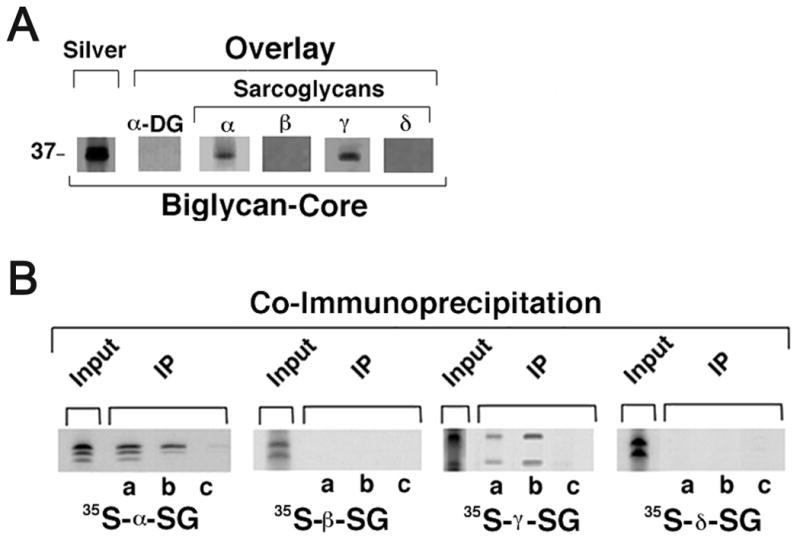
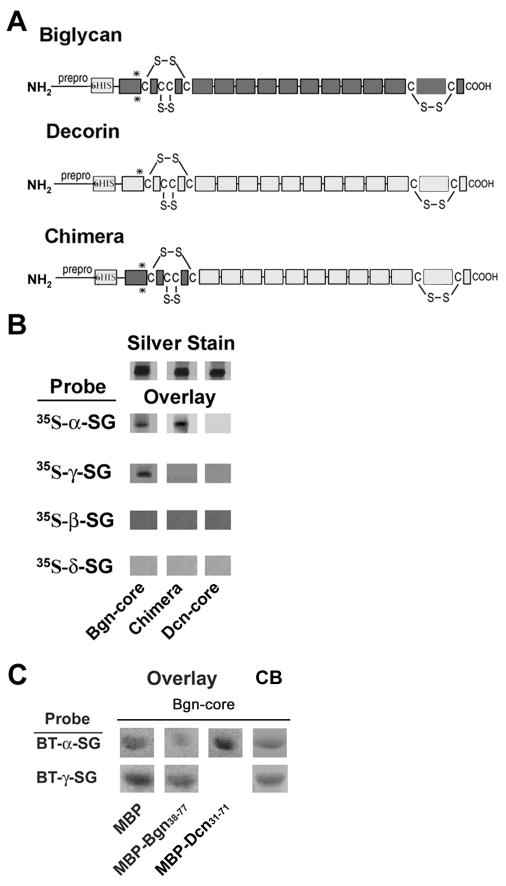
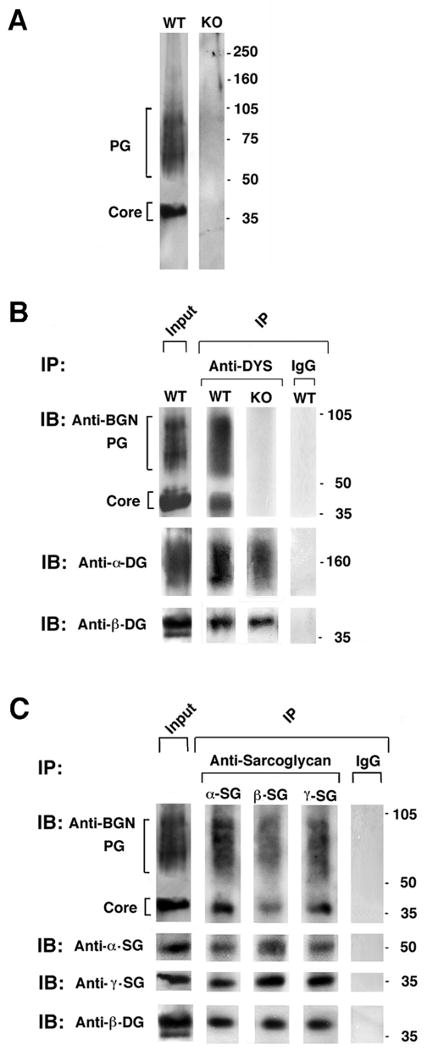
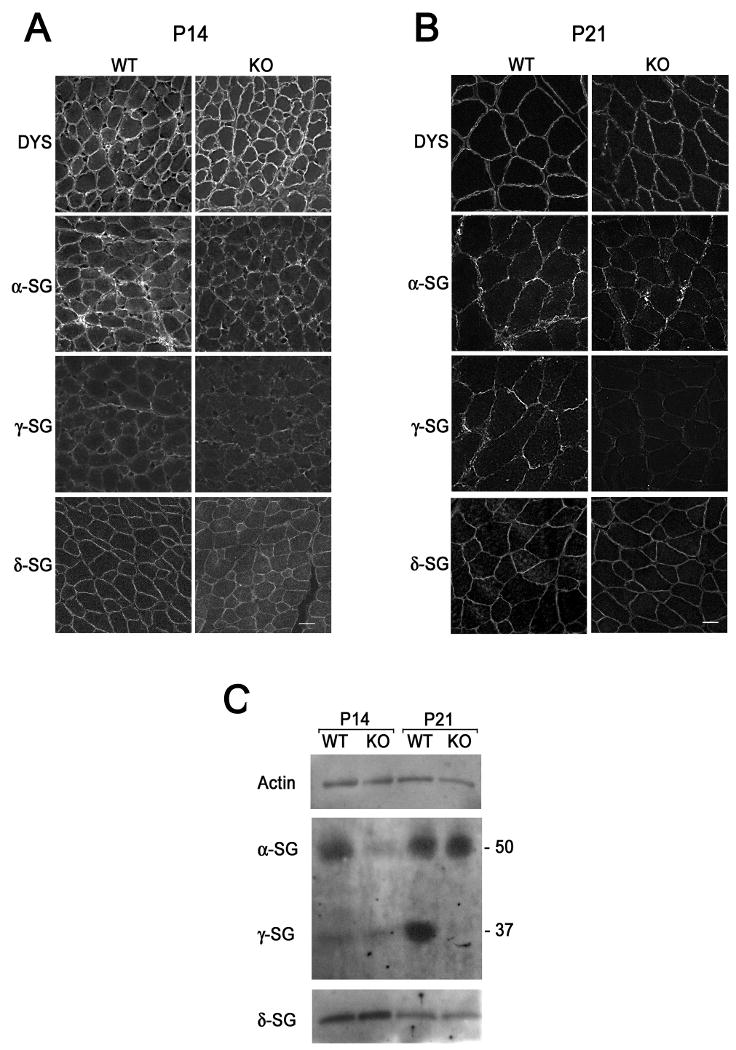
The dystrophin-associated protein complex (DAPC), which links the cytoskeleton to the extracellular matrix, is essential for muscle cell survival, and is defective in a wide range of muscular dystrophies. The DAPC contains two transmembrane subcomplexes-the dystroglycans and the sarcoglycans. Although several extracellular binding partners have been identified for the dystroglycans, none have been described for the sarcoglycan subcomplex. Here we show that the small leucine-rich repeat (LRR) proteoglycan biglycan binds to alpha- and gamma-sarcoglycan as judged by ligand blot overlay and co-immunoprecipitation assays. Our studies with biglycan-decorin chimeras show that alpha- and gamma-sarcoglycan bind to distinct sites on the polypeptide core of biglycan. Both biglycan proteoglycan as well as biglycan polypeptide lacking glycosaminoglycan (GAG) side chains are components of the dystrophin glycoprotein complex isolated from adult skeletal muscle membranes. Finally, our immunohistochemical and biochemical studies with biglycan null mice show that the expression of alpha- and gamma-sarcoglycan is selectively reduced in muscle from young (P14-P21) animals, while levels in adult muscle (> or = P35) are unchanged. We conclude that biglycan is a ligand for two members of the sarcoglycan complex and regulates their expression at discrete developmental ages.
Copyright 2006 Wiley-Liss, Inc.
Figures
References
-
- Adams ME, Butler MH, Dwyer TM, Peters MF, Murnane AA, Froehner SC. Two forms of mouse syntrophin, a 58 kd dystrophin-associated protein, differ in primary structure and tissue distribution. Neuron. 1993;11(3):531–540. - PubMed
-
- Ahn AH, Freener CA, Gussoni E, Yoshida M, Ozawa E, Kunkel LM. The three human syntrophin genes are expressed in diverse tissues, have distinct chromosomal locations, and each bind to dystrophin and its relatives. J Biol Chem. 1996;271(5):2724–2730. - PubMed
-
- Ameye L, Young MF. Mice deficient in small leucine-rich proteoglycans: novel in vivo models for osteoporosis, osteoarthritis, Ehlers-Danlos syndrome, muscular dystrophy, and corneal diseases. Glycobiology. 2002;12(9):107R–116R. - PubMed
-
- Bianco P, Fisher LW, Young MF, Termine JD, Robey PG. Expression and localization of the two small proteoglycans biglycan and decorin in developing human skeletal and non-skeletal tissues. J Histochem Cytochem. 1990;38(11):1549–1563. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
