

Comparison of rRNA and polar-lipid-derived fatty acid biomarkers for assessment of 13C-substrate incorporation by microorganisms in marine sediments
- PMID: 16885272
- PMCID: PMC1538766
- DOI: 10.1128/AEM.00423-06
Comparison of rRNA and polar-lipid-derived fatty acid biomarkers for assessment of 13C-substrate incorporation by microorganisms in marine sediments
Abstract
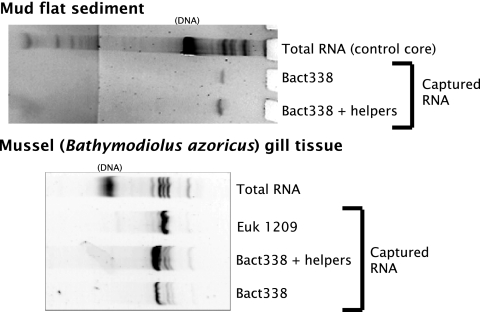
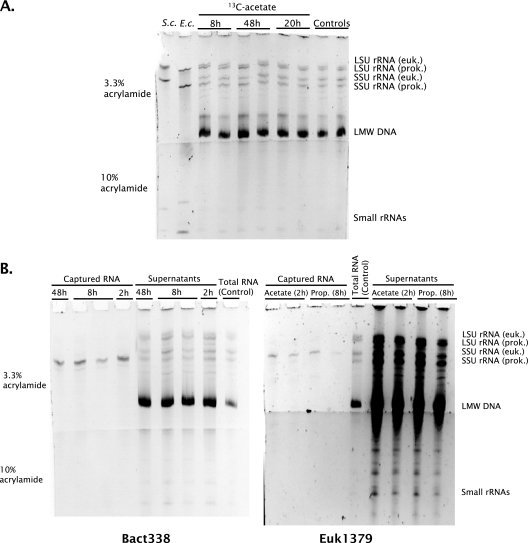
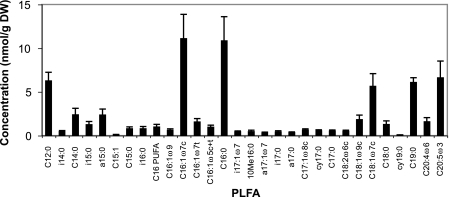
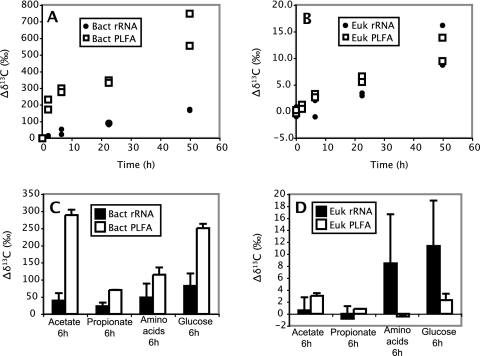
We determined whether a recently developed method to isolate specific small-subunit (SSU) rRNAs can be used in 13C-labeling studies to directly link community structure and function in natural ecosystems. Replicate North Sea sediment cores were incubated at the in situ temperature following addition of 13C-labeled acetate, propionate, amino acids, or glucose. Eukaryotic and bacterial SSU rRNAs were separated from total RNA by means of biotin-labeled oligonucleotide probes and streptavidin-coated paramagnetic beads, and the 13C content of the isolated rRNA was determined by elemental analysis-isotope ratio mass spectrometry. The SSU rRNA yield with the bead-capture protocol was improved by using helper probes. Incorporation of label into bacterial SSU rRNA was detectable after 2 h of incubation. The labeling was always much greater in bacterial SSU rRNA than in eukaryotic SSU rRNA, suggesting that bacteria were the main consumers of the 13C-labeled compounds. Similar results were obtained with the 13C-labeled polar-lipid-derived fatty acid (PLFA) approach, except that more label was detected in bacterial PLFA than in bacterial SSU rRNA. This may be attributable to the generally slow growth of sediment microbial populations, which results in low ribosome synthesis rates and relatively few ribosomes per cell. We discuss possible ways to improve the probe-capture protocol and the sensitivity of the 13C analysis of the captured SSU rRNA.
Figures


References
-
- Acimovic, Y., and I. R. Coe. 2002. Molecular evolution of the equilibrative nucleoside transporter family: identification of novel family members in prokaryotes and eukaryotes. Mol. Biol. Evol. 19:2199-2210. - PubMed
-
- Alm, E. W., and D. A. Stahl. 2000. Critical factors influencing the recovery and integrity of rRNA extracted from environmental samples: use of an optimized protocol to measure depth-related biomass distribution in freshwater sediments. J. Microbiol. Methods 40:153-162. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
