

A functional dlt operon, encoding proteins required for incorporation of d-alanine in teichoic acids in gram-positive bacteria, confers resistance to cationic antimicrobial peptides in Streptococcus pneumoniae
- PMID: 16885447
- PMCID: PMC1540085
- DOI: 10.1128/JB.00336-06
A functional dlt operon, encoding proteins required for incorporation of d-alanine in teichoic acids in gram-positive bacteria, confers resistance to cationic antimicrobial peptides in Streptococcus pneumoniae
Abstract
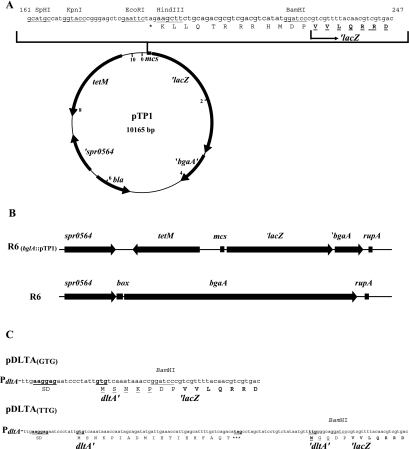
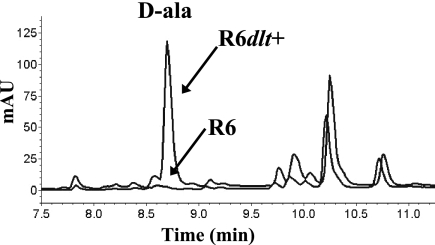
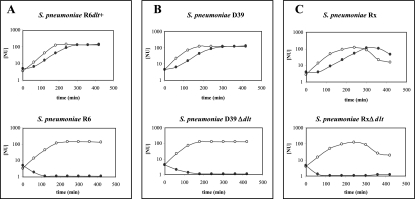
Streptococcus pneumoniae is one of the few species within the group of low-G +C gram-positive bacteria reported to contain no d-alanine in teichoic acids, although the dltABCD operon encoding proteins responsible for d-alanylation is present in the genomes of two S. pneumoniae strains, the laboratory strain R6 and the clinical isolate TIGR4. The annotation of dltA in R6 predicts a protein, d-alanine-d-alanyl carrier protein ligase (Dcl), that is shorter at the amino terminus than all other Dcl proteins. Translation of dltA could also start upstream of the annotated TTG start codon at a GTG, resulting in the premature termination of dltA translation at a stop codon. Applying a novel integrative translation probe plasmid with Escherichia coli 'lacZ as a reporter, we could demonstrate that dltA translation starts at the upstream GTG. Consequently, S. pneumoniae R6 is a dltA mutant, whereas S. pneumoniae D39, the parental strain of R6, and Rx, another derivative of D39, contained intact dltA genes. Repair of the stop codon in dltA of R6 and insertional inactivation of dltA in D39 and Rx yielded pairs of dltA-deficient and dltA-proficient strains. Subsequent phenotypic analysis showed that dltA inactivation resulted in enhanced sensitivity to the cationic antimicrobial peptides nisin and gallidermin, a phenotype fully consistent with those of dltA mutants of other gram-positive bacteria. In addition, mild alkaline hydrolysis of heat-inactivated whole cells released d-alanine from dltA-proficient strains, but not from dltA mutants. The results of our study suggest that, as in many other low-G+C gram-positive bacteria, teichoic acids of S. pneumoniae contain d-alanine residues in order to protect this human pathogen against the actions of cationic antimicrobial peptides.
Figures

References
-
- Abachin, E., C. Poyart, E. Pellegrini, E. Milohanic, F. Fiedler, P. Berche, and P. Trieu-Cuot. 2002. Formation of d-alanyl-lipoteichoic acid is required for adhesion and virulence of Listeria monocytogenes. Mol. Microbiol. 43:1-14. - PubMed
-
- Argaman, L., R. Hershberg, J. Vogel, G. Bejerano, E. G. Wagner, H. Margalit, and S. Altuvia. 2001. Novel small RNA-encoding genes in the intergenic regions of Escherichia coli. Curr. Biol. 11:941-950. - PubMed
-
- Behr, T., W. Fischer, J. Peter-Katalinic, and H. Egge. 1992. The structure of pneumococcal lipoteichoic acid. Improved preparation, chemical and mass spectrometric studies. Eur. J. Biochem. 207:1063-1075. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
