

Proteinuria precedes podocyte abnormalities inLamb2-/- mice, implicating the glomerular basement membrane as an albumin barrier
- PMID: 16886065
- PMCID: PMC1523402
- DOI: 10.1172/JCI28414
Proteinuria precedes podocyte abnormalities inLamb2-/- mice, implicating the glomerular basement membrane as an albumin barrier
Abstract
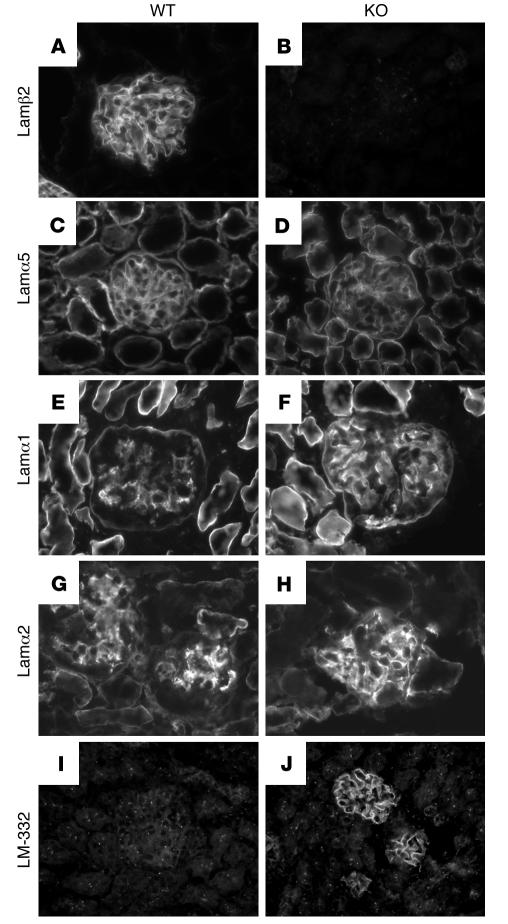
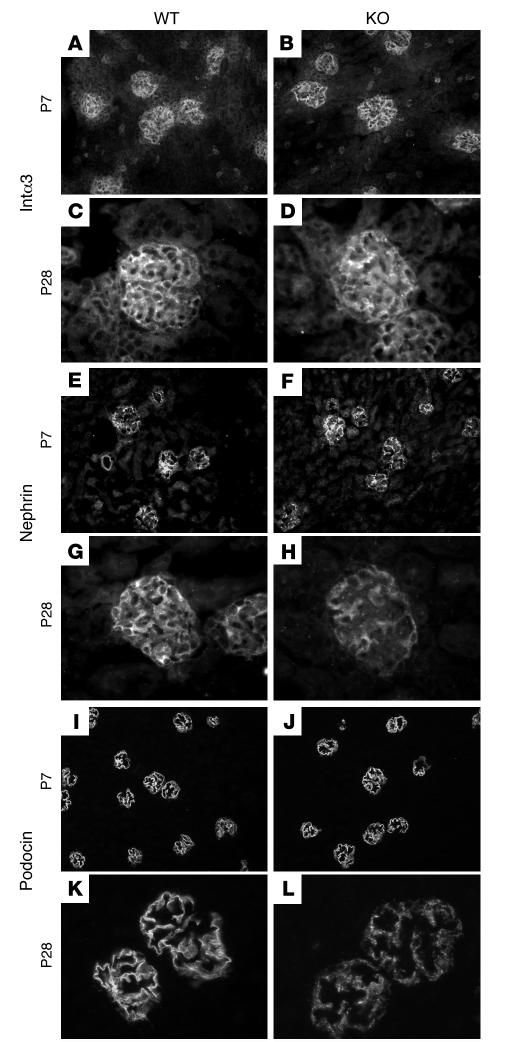
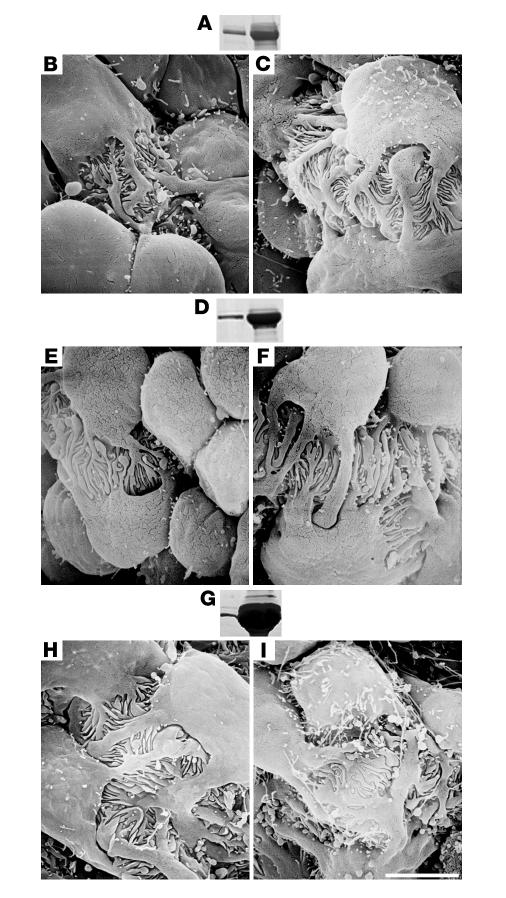
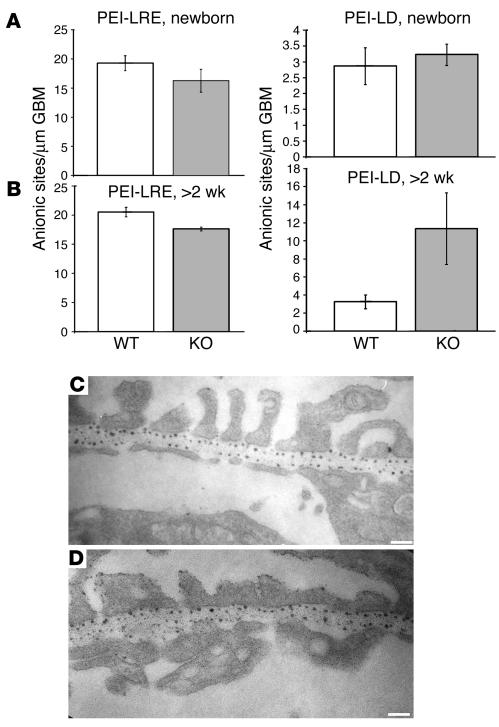
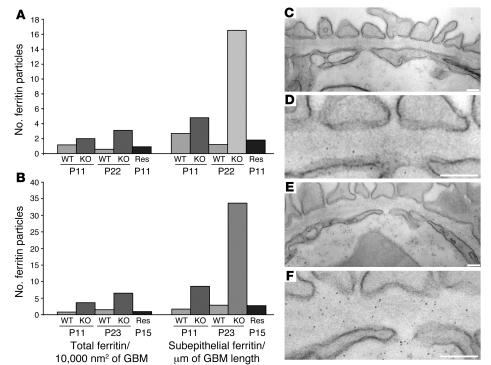
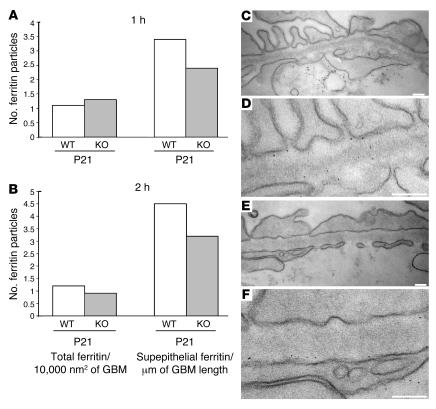
Primary defects in either podocytes or the glomerular basement membrane (GBM) cause proteinuria, a fact that complicates defining the barrier to albumin. Laminin beta2 (LAMB2) is a GBM component required for proper functioning of the glomerular filtration barrier. To investigate the GBM's role in glomerular filtration, we characterized GBM and overlying podocyte architecture in relation to development and progression of proteinuria in Lamb2-/- mice, which model Pierson syndrome, a rare congenital nephrotic syndrome. We found ectopic deposition of several laminins and mislocalization of anionic sites in the GBM, which together suggest that the Lamb2-/- GBM is severely disorganized, although it is ultrastructurally intact. Importantly, albuminuria was detectable shortly after birth and preceded podocyte foot process effacement and loss of slit diaphragms by at least 7 days. Expression and localization of slit diaphragm and foot process-associated proteins appeared normal at early stages. GBM permeability to the electron-dense tracer ferritin was dramatically elevated in Lamb2-/- mice, even before widespread foot process effacement. Increased ferritin permeability was not observed in nephrotic CD2-associated protein-null (Cd2ap-/-) mice, which have a primary podocyte defect. Together these data show that the GBM serves as a barrier to protein in vivo and that the glomerular slit diaphragm alone is not sufficient to prevent the passage of albumin into the urinary space.
Figures
Comment in
-
The glomerular basement membrane: not gone, just forgotten.J Clin Invest. 2006 Aug;116(8):2090-3. doi: 10.1172/JCI29488. J Clin Invest. 2006. PMID: 16886057 Free PMC article.
References
-
- Miner J.H., Yurchenco P.D. Laminin functions in tissue morphogenesis. Annu. Rev. Cell Dev. Biol. 2004;20:255–284. - PubMed
-
- Miner J.H. Renal basement membrane components. Kidney Int. 1999;56:2016–2024. - PubMed
-
- Aumailley M., et al. A simplified laminin nomenclature. Matrix Biol. 2005;24:326–332. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
