

Diverse immunoglobulin light chain organizations in fish retain potential to revise B cell receptor specificities
- PMID: 16888007
- PMCID: PMC3129705
- DOI: 10.4049/jimmunol.177.4.2452
Diverse immunoglobulin light chain organizations in fish retain potential to revise B cell receptor specificities
Abstract
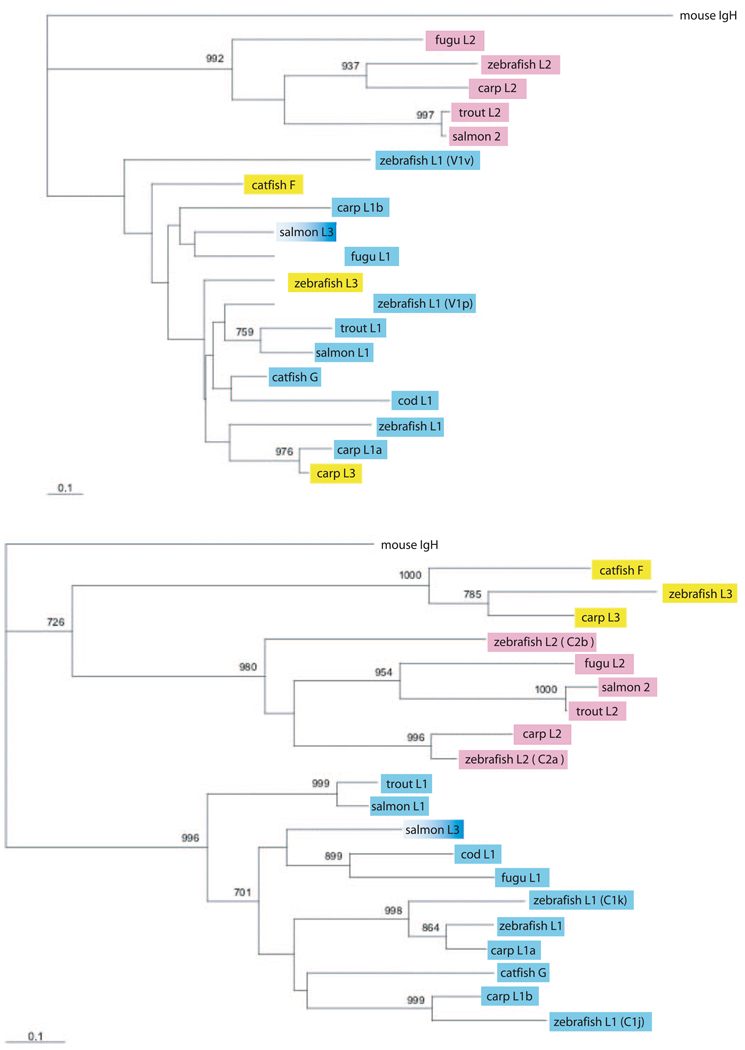
We have characterized the genomic organization of the three zebrafish L chain isotypes and found they all differed from those reported in other teleost fishes. Two of the zebrafish L chain isotypes are encoded by two loci, each carrying multiple V gene segments. To understand the derivation of these L chain genes and their organizations, we performed phylogenetic analyses and show that IgL organization can diverge considerably among closely related species. Except in zebrafish, the teleost fish IgL each contain only two to four recombinogenic components (one to three V, one J) and exist in multiple copies. BCR heterogeneity can be generated, but this arrangement apparently provides neither combinatorial diversification nor an opportunity for the secondary rearrangements that, in mammals, take place during receptor editing, a process crucial to the promotion of tolerance in developing lymphocytes. Examination of the zebrafish IgL recombination possibilities gave insight into how the suppression of self-reactivity by receptor editing might be managed, including in miniloci. We suggest that, despite the diverse IgL organizations in early and higher vertebrates, two elements essential to generating the Ab repertoire are retained: the numerous genes/loci for ligand-binding diversification and the potential for correcting unwanted specificities that arise.
Conflict of interest statement
The authors have no financial conflict of interest.
Figures




References
-
- Riblet R. Immunoglobulin heavy chain genes of mouse. In: Honjo T, Alt FW, Neuberger MS, editors. Molecular Biology of B Cells. Amsterdam: Elsevier Academic Press; 2004. pp. 19–26.
-
- Fugmann SD, Lee AI, Shockett PE, Villey IJ, Schatz DG. The RAG proteins and V(D)J recombination: complexes, ends, and transposition. Annu. Rev. Immunol. 2000;18:495–527. - PubMed
-
- Zachau HG. Immunoglobulin κ genes in human and mouse. In: Honjo T, Alt FW, Neuberger MS, editors. Molecular Biology of B Cells. Amsterdam: Elsevier Academic Press; 2004. pp. 27–36.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
