

IMMUTANS does not act as a stress-induced safety valve in the protection of the photosynthetic apparatus of Arabidopsis during steady-state photosynthesis
- PMID: 16891546
- PMCID: PMC1586030
- DOI: 10.1104/pp.106.085886
IMMUTANS does not act as a stress-induced safety valve in the protection of the photosynthetic apparatus of Arabidopsis during steady-state photosynthesis
Abstract
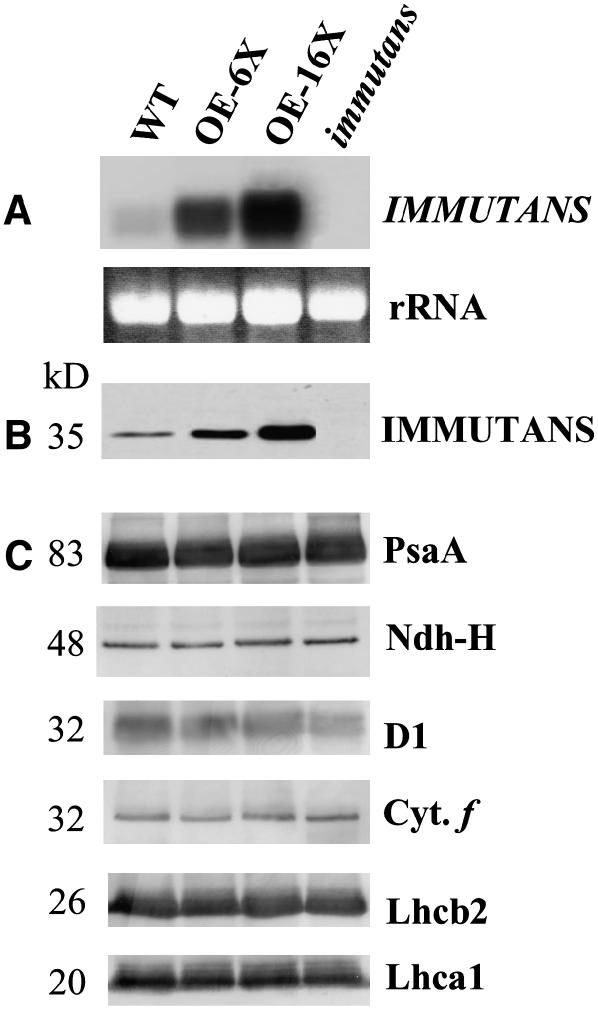
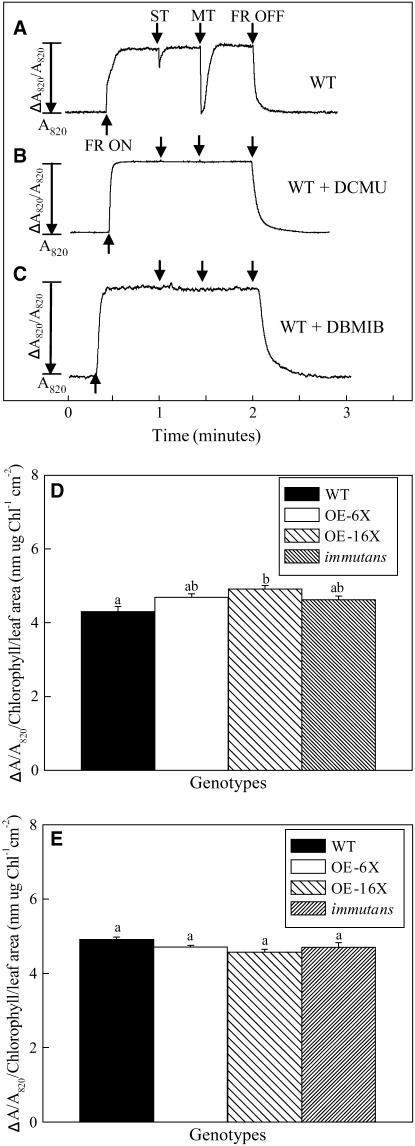
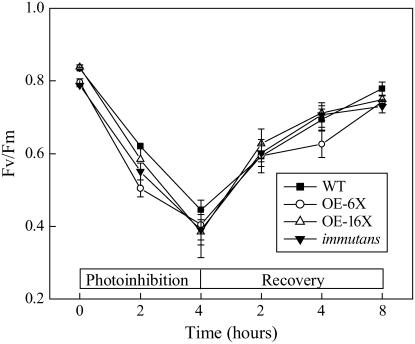
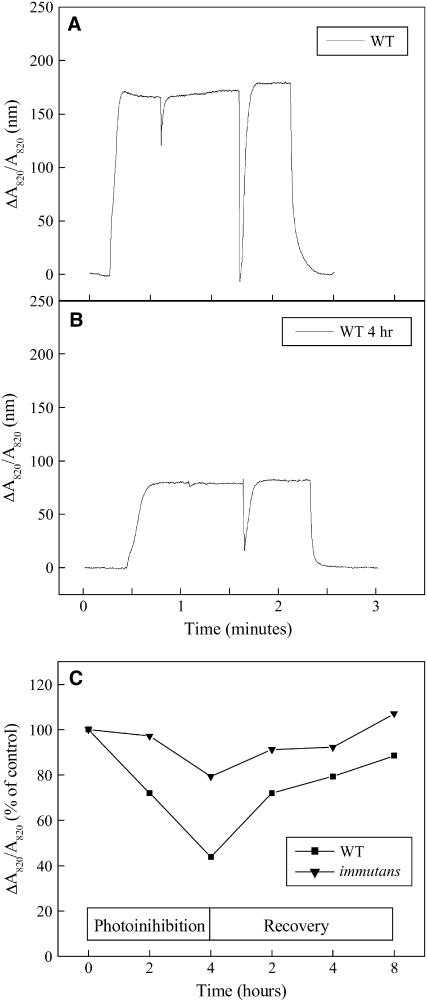
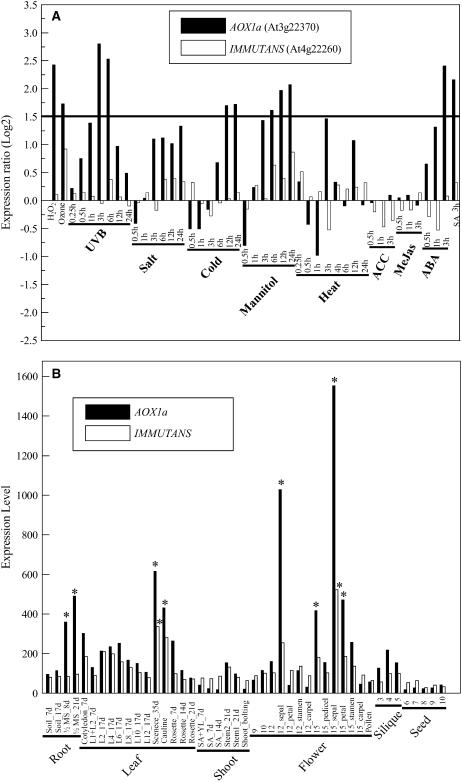
IMMUTANS (IM) encodes a thylakoid membrane protein that has been hypothesized to act as a terminal oxidase that couples the reduction of O(2) to the oxidation of the plastoquinone (PQ) pool of the photosynthetic electron transport chain. Because IM shares sequence similarity to the stress-induced mitochondrial alternative oxidase (AOX), it has been suggested that the protein encoded by IM acts as a safety valve during the generation of excess photosynthetically generated electrons. We combined in vivo chlorophyll fluorescence quenching analyses with measurements of the redox state of P(700) to assess the capacity of IM to compete with photosystem I for intersystem electrons during steady-state photosynthesis in Arabidopsis (Arabidopsis thaliana). Comparisons were made between wild-type plants, im mutant plants, as well as transgenics in which IM protein levels had been overexpressed six (OE-6 x) and 16 (OE-16 x) times. Immunoblots indicated that IM abundance was the only major variant that we could detect between these genotypes. Overexpression of IM did not result in increased capacity to keep the PQ pool oxidized compared to either the wild type or im grown under control conditions (25 degrees C and photosynthetic photon flux density of 150 micromol photons m(-2) s(-1)). Similar results were observed either after 3-d cold stress at 5 degrees C or after full-leaf expansion at 5 degrees C and photosynthetic photon flux density of 150 micromol photons m(-2) s(-1). Furthermore, IM abundance did not enhance protection of either photosystem II or photosystem I from photoinhibition at either 25 degrees C or 5 degrees C. Our in vivo data indicate that modulation of IM expression and polypeptide accumulation does not alter the flux of intersystem electrons to P(700)(+) during steady-state photosynthesis and does not provide any significant photoprotection. In contrast to AOX1a, meta-analyses of published Arabidopsis microarray data indicated that IM expression exhibited minimal modulation in response to myriad abiotic stresses, which is consistent with our functional data. However, IM exhibited significant modulation in response to development in concert with changes in AOX1a expression. Thus, neither our functional analyses of the IM knockout and overexpression lines nor meta-analyses of gene expression support the model that IM acts as a safety valve to regulate the redox state of the PQ pool during stress and acclimation. Rather, IM appears to be strongly regulated by developmental stage of Arabidopsis.
Figures


References
-
- Aluru MR, Rodermel SR (2004) Control of chloroplast redox by the IMMUTANS terminal oxidase. Physiol Plant 120: 4–11 - PubMed
-
- Aluru MR, Yu F, Fu A, Rodermel SR (2006) Arabidopsis variegation mutants: new insights into chloroplast biogenesis. J Exp Bot 57: 1871-1881 - PubMed
-
- Asada K, Heber U, Schreiber U (1993) Electron flow to the intersystem chain from stromal components and cyclic electron flow in maize chloroplasts, as detected in intact leaves by monitoring redox change of P700 and chlorophyll fluorescence. Plant Cell Physiol 34: 39–50
-
- Baerr JN, Thomas JD, Taylor BG, Rodermel SR, Gray GR (2005) Differential photosynthetic compensatory mechanisms exist in the immutans mutant of Arabidopsis thaliana. Physiol Plant 124: 390–402
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
