

Molecular and functional analysis of nicotinate catabolism in Eubacterium barkeri
- PMID: 16894175
- PMCID: PMC1562527
- DOI: 10.1073/pnas.0601635103
Molecular and functional analysis of nicotinate catabolism in Eubacterium barkeri
Abstract
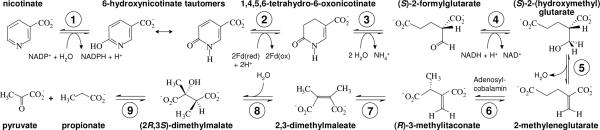
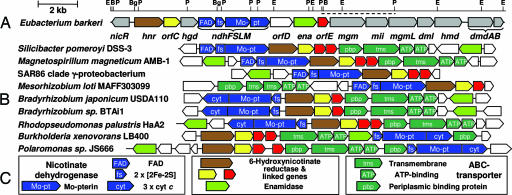
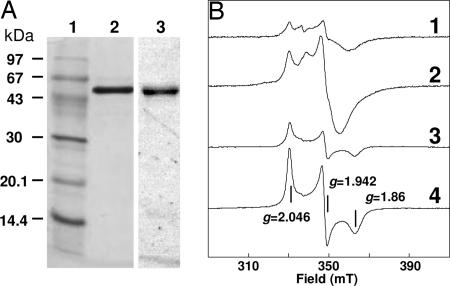
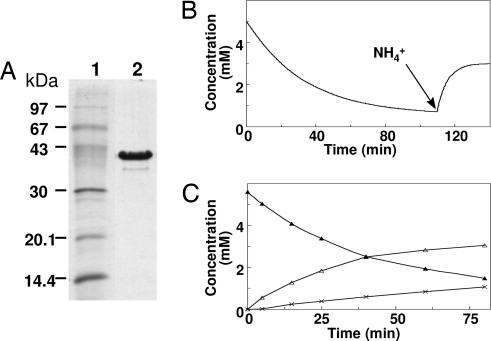
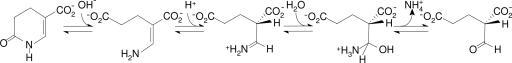
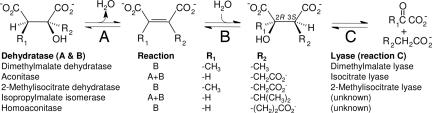
The anaerobic soil bacterium Eubacterium barkeri catabolizes nicotinate to pyruvate and propionate via a unique fermentation. A full molecular characterization of nicotinate fermentation in this organism was accomplished by the following results: (i) A 23.2-kb DNA segment with a gene cluster encoding all nine enzymes was cloned and sequenced, (ii) two chiral intermediates were discovered, and (iii) three enzymes were found, completing the hitherto unknown part of the pathway. Nicotinate dehydrogenase, a (nonselenocysteine) selenium-containing four-subunit enzyme, is encoded by ndhF (FAD subunit), ndhS (2 x [2Fe-2S] subunit), and by the ndhL/ndhM genes. In contrast to all enzymes of the xanthine dehydrogenase family, the latter two encode a two-subunit molybdopterin protein. The 6-hydroxynicotinate reductase, catalyzing reduction of 6-hydroxynicotinate to 1,4,5,6-tetrahydro-6-oxonicotinate, was purified and shown to contain a covalently bound flavin cofactor, one [2Fe-2S](2+/1+) and two [4Fe-4S](2+/1+) clusters. Enamidase, a bifunctional Fe-Zn enzyme belonging to the amidohydrolase family, mediates hydrolysis of 1,4,5,6-tetrahydro-6-oxonicotinate to ammonia and (S)-2-formylglutarate. NADH-dependent reduction of the latter to (S)-2-(hydroxymethyl)glutarate is catalyzed by a member of the 3-hydroxyisobutyrate/phosphogluconate dehydrogenase family. A [4Fe-4S]-containing serine dehydratase-like enzyme is predicted to form 2-methyleneglutarate. After the action of the coenzyme B(12)-dependent 2-methyleneglutarate mutase and 3-methylitaconate isomerase, an aconitase and isocitrate lyase family pair of enzymes, (2R,3S)-dimethylmalate dehydratase and lyase, completes the pathway. Genes corresponding to the first three enzymes of the E. barkeri nicotinate catabolism were identified in nine Proteobacteria.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures
References
-
- London J., Knight M. J. Gen. Microbiol. 1966;44:241–254. - PubMed
-
- Andreesen J. R., Fetzner S. Met. Ions Biol. Syst. 2002;39:405–430. - PubMed
-
- Nakano H., Wieser M., Hurh B., Kawai T., Yoshida T., Yamane T., Nagasawa T. Eur. J. Biochem. 1999;260:120–126. - PubMed
-
- Nagel M., Andreesen J. R. Arch. Microbiol. 1990;154:605–613.
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
