

Endogenous bone morphogenetic protein antagonists regulate mammalian neural crest generation and survival
- PMID: 16894609
- PMCID: PMC6626635
- DOI: 10.1002/dvdy.20891
Endogenous bone morphogenetic protein antagonists regulate mammalian neural crest generation and survival
Abstract
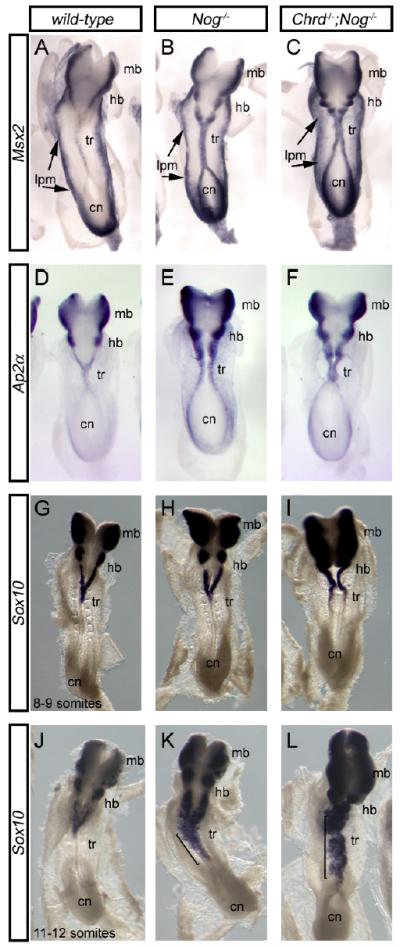
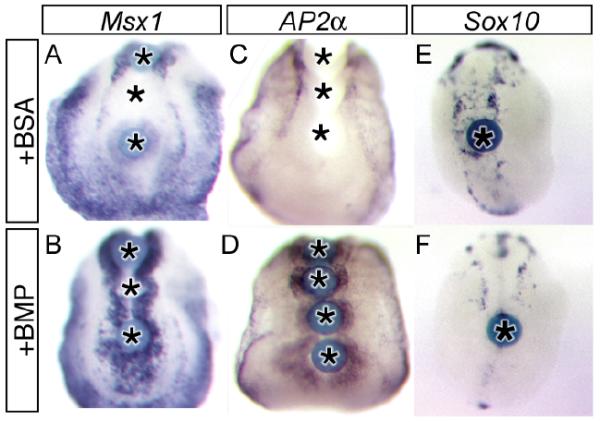
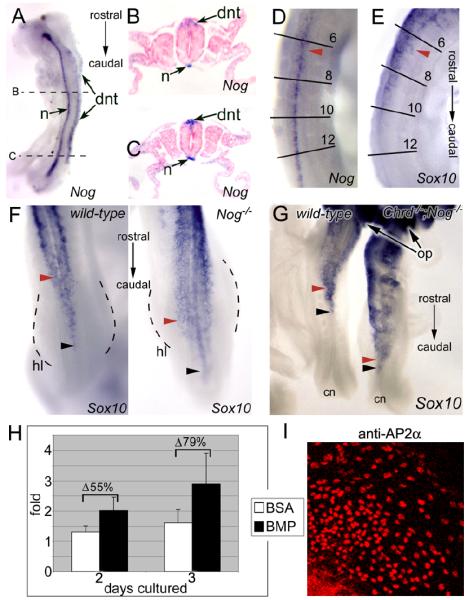
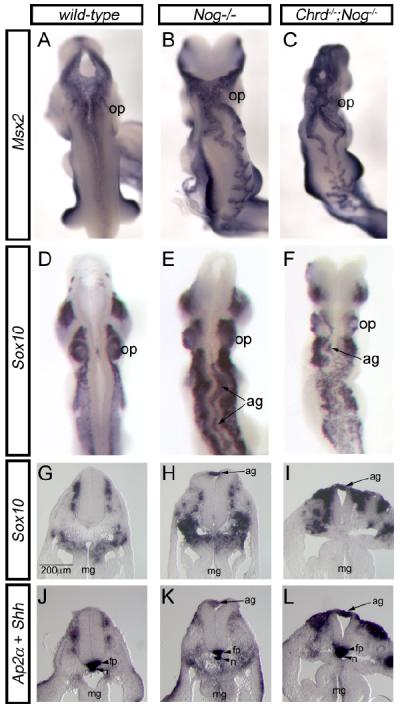
We demonstrate here that Chordin and Noggin function as bone morphogenetic protein (BMP) antagonists in vivo to promote mammalian neural crest development. Using Chrd and Nog single and compound mutants, we find that Noggin has a major role in promoting neural crest formation, in which Chordin is partially redundant. BMP signaling is increased in dorsal tissues lacking Noggin and is further increased when Chordin is also absent. The early neural crest domain is expanded with decreased BMP antagonism in vivo. Noggin and Chordin also regulate subsequent neural crest cell emigration from the neural tube. However, reduced levels of these BMP antagonists ultimately result in perturbation of neural crest cell derived peripheral nervous system and craniofacial skeletal elements. Such defects reflect, at least in part, a function to limit apoptosis in neural crest cells. Noggin and Chordin, therefore, function together to regulate both the generation and survival of neural crest cells in mammalian development.
Copyright 2006 Wiley-Liss, Inc.
Figures




References
-
- Abzhanov A, Protas M, Grant BR, Grant PR, Tabin CJ. Bmp4 and morphological variation of beaks in Darwin’s finches. Science. 2004;305:1464–1465. - PubMed
-
- Abu-Issa R, Smyth G, Smoak I, Yamamura K, Meyers EN. Fgf8 is required for pharyngeal arch and cardiovascular development in the mouse. Development. 2002;129:4613–25. - PubMed
-
- Ahlgren SC, Bronner-Fraser M. Inhibition of sonic hedgehog signaling in vivo results in craniofacial neural crest cell death. Curr Biol. 1999;9:1304–14. - PubMed
-
- Anderson RM, Lawrence AR, Stottmann RW, Bachiller D, Klingensmith J. Chordin and noggin promote organizing centers of forebrain development in the mouse. Development. 2002;129:4975–87. - PubMed
-
- Arkell R, Beddington RSP. BMP-7 influences pattern and growth of the developing hindbrain of mouse embryos. Devlopment. 1997;124:1–12. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
