

Binding of myotrophin/V-1 to actin-capping protein: implications for how capping protein binds to the filament barbed end
- PMID: 16895918
- PMCID: PMC2277501
- DOI: 10.1074/jbc.M606278200
Binding of myotrophin/V-1 to actin-capping protein: implications for how capping protein binds to the filament barbed end
Abstract
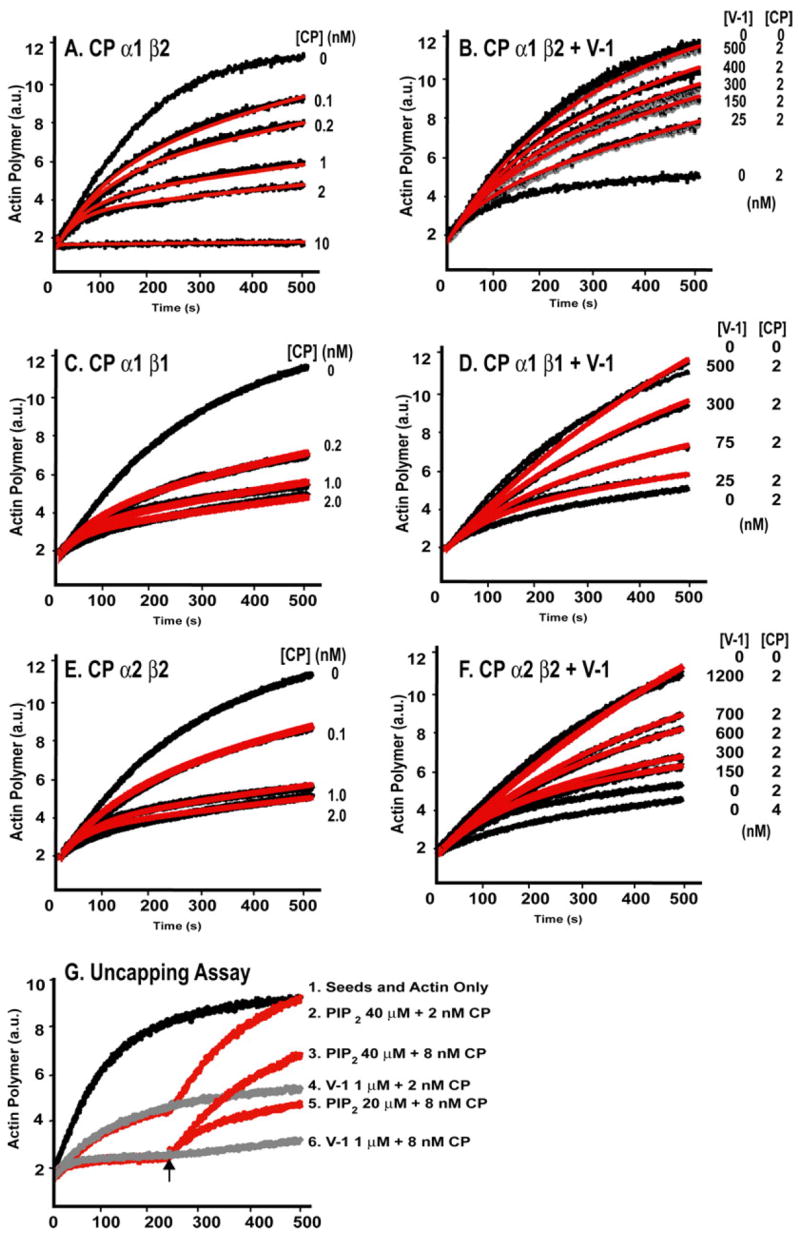
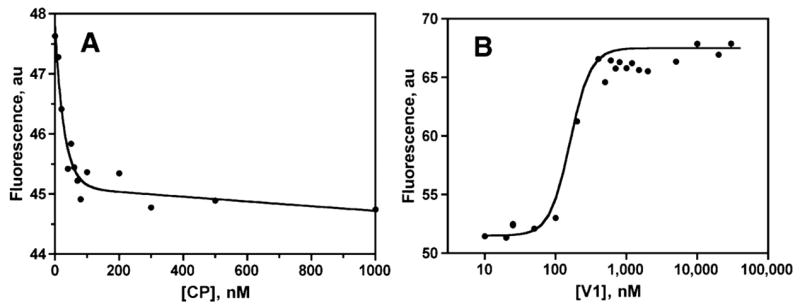
The heterodimeric actin-capping protein (CP) regulates actin assembly and cell motility by binding tightly to the barbed end of the actin filament. Here we demonstrate that myotrophin/V-1 binds directly to CP in a 1:1 molar ratio with a Kd of 10-50 nm. V-1 binding inhibited the ability of CP to cap the barbed ends of actin filaments. The actin-binding COOH-terminal region, the "tentacle," of the CP beta subunit was important for binding V-1, with lesser contributions from the alpha subunit COOH-terminal region and the body of the protein. V-1 appears to be unable to bind to CP that is on the barbed end, based on the observations that V-1 had no activity in an uncapping assay and that the V-1.CP complex had no capping activity. Two loops of V-1, which extend out from the alpha-helical backbone of this ankyrin repeat protein, were necessary for V-1 to bind CP. Parallel computational studies determined a bound conformation of the beta tentacle with V-1 that is consistent with these findings, and they offered insight into experimentally observed differences between the alpha1 and alpha2 isoforms as well as the mutant lacking the alpha tentacle. These results support and extend our "wobble" model for CP binding to the actin filament, in which the two COOH-terminal regions of CP bind independently to the actin filament, and bound CP is able to wobble when attached only via its mobile beta-subunit tentacle. This model is also supported by molecular dynamics simulations of CP reported here. The existence of the wobble state may be important for actin dynamics in cells.
Figures







References
-
- Sen S, Kundu G, Mekhail N, Castel J, Misono K, Healy B. J Biol Chem. 1990;27:16635–16643. - PubMed
-
- Sil P, Misono K, Sen S. Circ Res. 1993;1:98–108. - PubMed
-
- Sarkar S, Leaman DW, Gupta S, Sil P, Young D, Morehead A, Mukherjee D, Ratliff N, Sun Y, Rayborn M, Hollyfield J, Sen S. J Biol Chem. 2004;19:20422–20434. - PubMed
-
- Pennica D, Shaw KJ, Luoh SM, Wood WI. Gene (Amst) 1995;158:305–306. - PubMed
-
- Taoka M, Isobe T, Okuyama T, Watanabe M, Kondo H, Yamakawa Y, Ozawa F, Hishinuma F, Kubota M, Minegishi A. J Biol Chem. 1994;269:9946–9951. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
