

The vtc4 gene influences polyphosphate storage, morphogenesis, and virulence in the maize pathogen Ustilago maydis
- PMID: 16896223
- PMCID: PMC1539147
- DOI: 10.1128/EC.00131-06
The vtc4 gene influences polyphosphate storage, morphogenesis, and virulence in the maize pathogen Ustilago maydis
Abstract
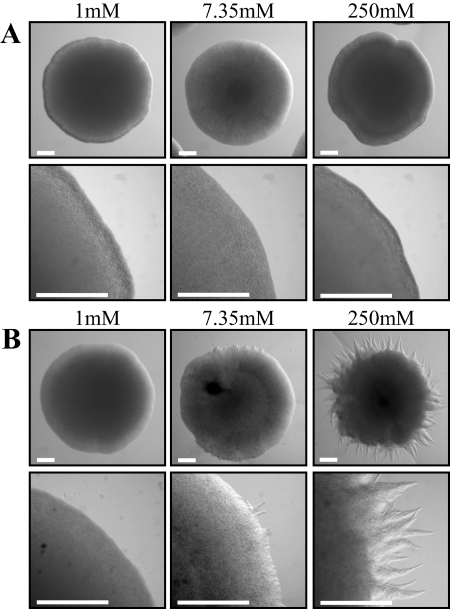
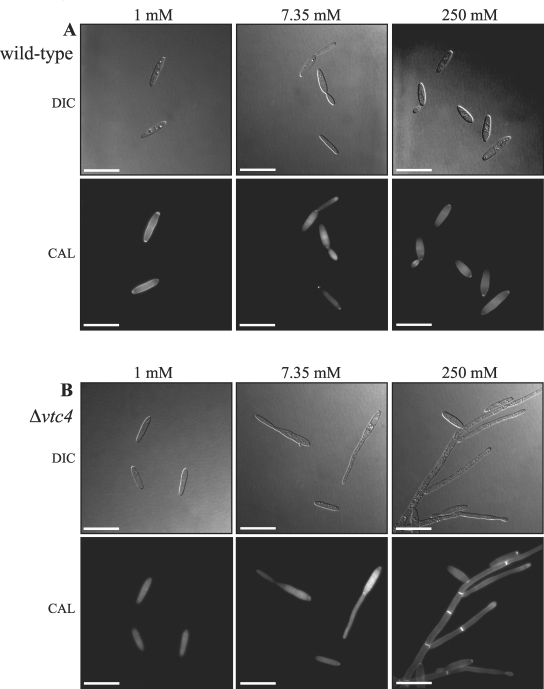
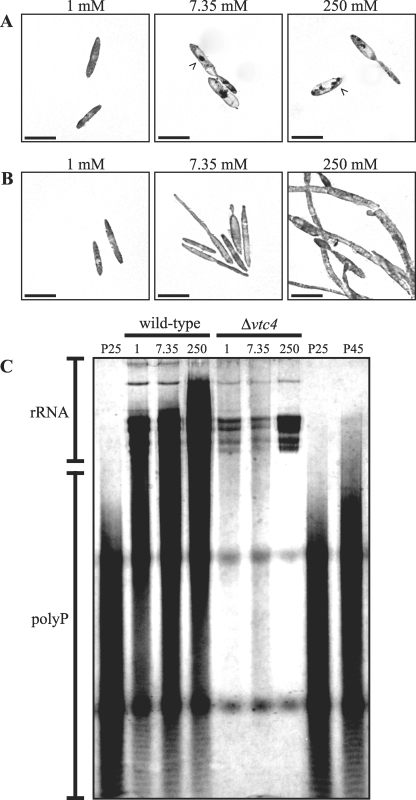
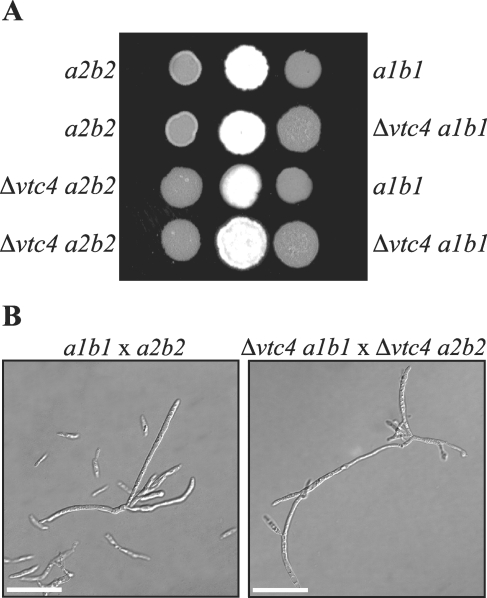
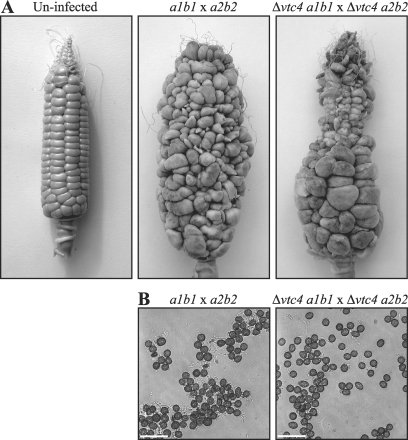
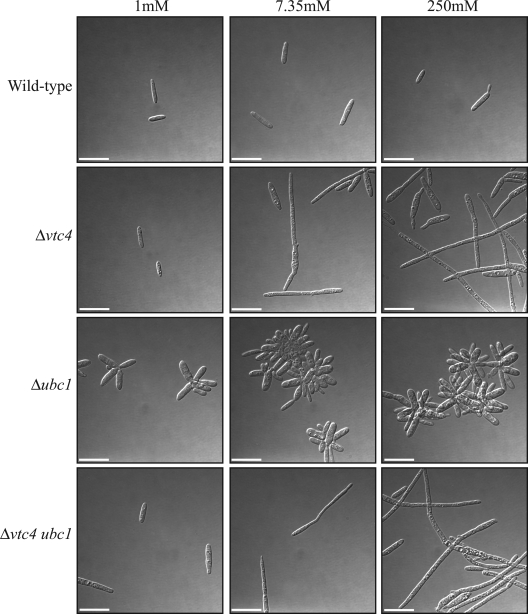
The maize pathogen Ustilago maydis switches from budding to filamentous, dikaryotic growth in response to environmental signals including nutrient status, growth in the host, and the presence of mating pheromones. The filamentous dikaryon is capable of proliferating within host tissue to cause disease symptoms including tumors. The transition from yeast cells to hyphal filaments is regulated by a mitogen-activated protein kinase cascade and a cyclic-AMP-protein kinase A (PKA) pathway. Serial analysis of gene expression with PKA mutants identified orthologs of components of the PHO phosphate acquisition pathway as transcriptional targets of the PKA pathway, and these included genes for Pho84, an acid phosphatase, and the vacuolar transport chaperones Vtc1 and Vtc4. In Saccharomyces cerevisiae, Vtc4p is required during the fusion of inorganic-phosphate-containing vesicles to the vacuolar membrane and the consequent accumulation of phosphate stored as polyphosphate (polyP) in the vacuole. We found that deletion of vtc4 in U. maydis also reduced polyP stored in vacuoles. Intriguingly, Deltavtc4 mutants possessed a filamentous cellular morphology, in contrast to the budding, yeast-like growth of the wild-type parent. The Deltavtc4 mutants also displayed decreased symptom development and reduced proliferation in planta. The interaction with PKA signaling was further investigated by the generation of Deltavtc4 ubc1 double mutants. Deletion of vtc4 completely suppressed the multiple-budded phenotype of a Deltaubc1 mutant, indicating that polyP stores are essential for this PKA-induced trait. Overall, this study reveals a novel role for PKA-regulated polyP accumulation in the control of fungal morphogenesis and virulence.
Figures
Similar articles
-
An Ustilago maydis septin is required for filamentous growth in culture and for full symptom development on maize.Eukaryot Cell. 2005 Dec;4(12):2044-56. doi: 10.1128/EC.4.12.2044-2056.2005. Eukaryot Cell. 2005. PMID: 16339722 Free PMC article.
-
Serial analysis of gene expression reveals conserved links between protein kinase A, ribosome biogenesis, and phosphate metabolism in Ustilago maydis.Eukaryot Cell. 2005 Dec;4(12):2029-43. doi: 10.1128/EC.4.12.2029-2043.2005. Eukaryot Cell. 2005. PMID: 16339721 Free PMC article.
-
Deletion of the Ustilago maydis ortholog of the Aspergillus sporulation regulator medA affects mating and virulence through pheromone response.Fungal Genet Biol. 2012 Jun;49(6):426-32. doi: 10.1016/j.fgb.2012.04.002. Epub 2012 Apr 19. Fungal Genet Biol. 2012. PMID: 22537792
-
Control of mating and development in Ustilago maydis.Antonie Van Leeuwenhoek. 1994;65(3):191-7. doi: 10.1007/BF00871946. Antonie Van Leeuwenhoek. 1994. PMID: 7847885 Review.
-
Genetics of Ustilago maydis, a fungal pathogen that induces tumors in maize.Annu Rev Genet. 1995;29:179-208. doi: 10.1146/annurev.ge.29.120195.001143. Annu Rev Genet. 1995. PMID: 8825473 Review.
Cited by
-
Blocking Polyphosphate Mobilization Inhibits Pho4 Activation and Virulence in the Pathogen Candida albicans.mBio. 2022 Jun 28;13(3):e0034222. doi: 10.1128/mbio.00342-22. Epub 2022 May 16. mBio. 2022. PMID: 35575514 Free PMC article.
-
Polyphosphatases have a polyphosphate-independent influence on the virulence of Cryptococcus neoformans.Infect Immun. 2025 Apr 8;93(4):e0007225. doi: 10.1128/iai.00072-25. Epub 2025 Mar 12. Infect Immun. 2025. PMID: 40071953 Free PMC article.
-
Phosphate limitation enhances malic acid production on nitrogen-rich molasses with Ustilago trichophora.Biotechnol Biofuels Bioprod. 2024 Jul 3;17(1):92. doi: 10.1186/s13068-024-02543-z. Biotechnol Biofuels Bioprod. 2024. PMID: 38961457 Free PMC article.
-
Molecular characterization of CHAD domains as inorganic polyphosphate-binding modules.Life Sci Alliance. 2019 May 27;2(3):e201900385. doi: 10.26508/lsa.201900385. Print 2019 Jun. Life Sci Alliance. 2019. PMID: 31133615 Free PMC article.
-
The putative phospholipase Lip2 counteracts oxidative damage and influences the virulence of Ustilago maydis.Mol Plant Pathol. 2017 Feb;18(2):210-221. doi: 10.1111/mpp.12391. Epub 2016 Apr 21. Mol Plant Pathol. 2017. PMID: 26950180 Free PMC article.
References
-
- Andrews, D. L., J. D. Egan, M. E. Mayorga, and S. E. Gold. 2000. The Ustilago maydis ubc4 and ubc5 genes encode members of a MAP kinase cascade required for filamentous growth. Mol. Plant-Microbe Interact. 13:781-786. - PubMed
-
- Banuett, F., and I. Herskowitz. 1994. Identification of fuz7, a Ustilago maydis MEK/MAPKK homolog required for a-locus-dependent and -independent steps in the fungal life cycle. Genes Dev. 8:1367-1378. - PubMed
-
- Banuett, F., and I. Herskowitz. 1996. Discrete developmental stages during teliospore formation in the corn smut fungus, Ustilago maydis. Development 122:2965-2976. - PubMed
-
- Barrett, K. J., S. E. Gold, and J. W. Kronstad. 1993. Identification and complementation of a mutation to constitutive filamentous growth in Ustilago maydis. Mol. Plant-Microbe Interact. 6:274-283. - PubMed
-
- Bolker, M., S. Genin, C. Lehmler, and R. Kahmann. 1995. Genetic regulation of mating and dimorphism in Ustilago maydis. Can. J. Bot. 73:320-325.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
