

Laforin, a dual specificity phosphatase that dephosphorylates complex carbohydrates
- PMID: 16901901
- PMCID: PMC2774450
- DOI: 10.1074/jbc.M606117200
Laforin, a dual specificity phosphatase that dephosphorylates complex carbohydrates
Abstract
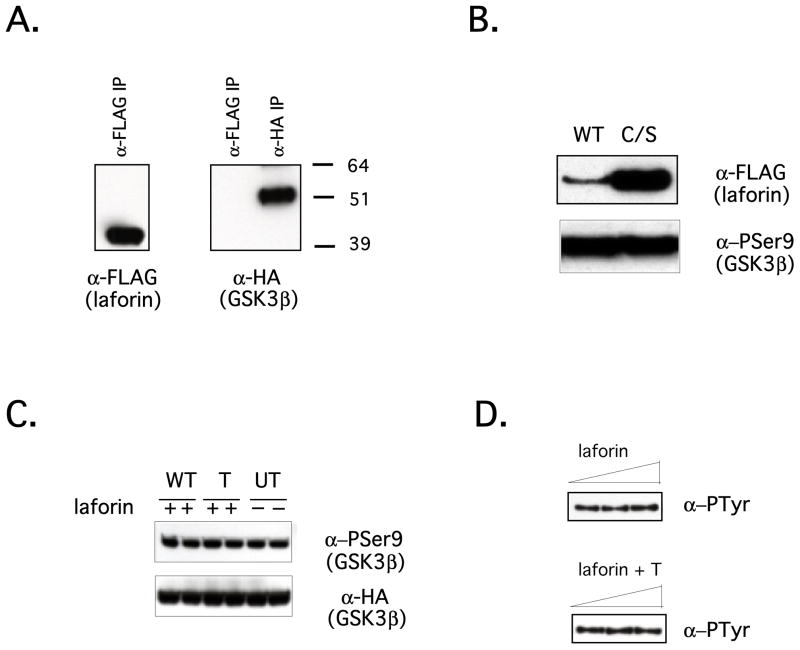
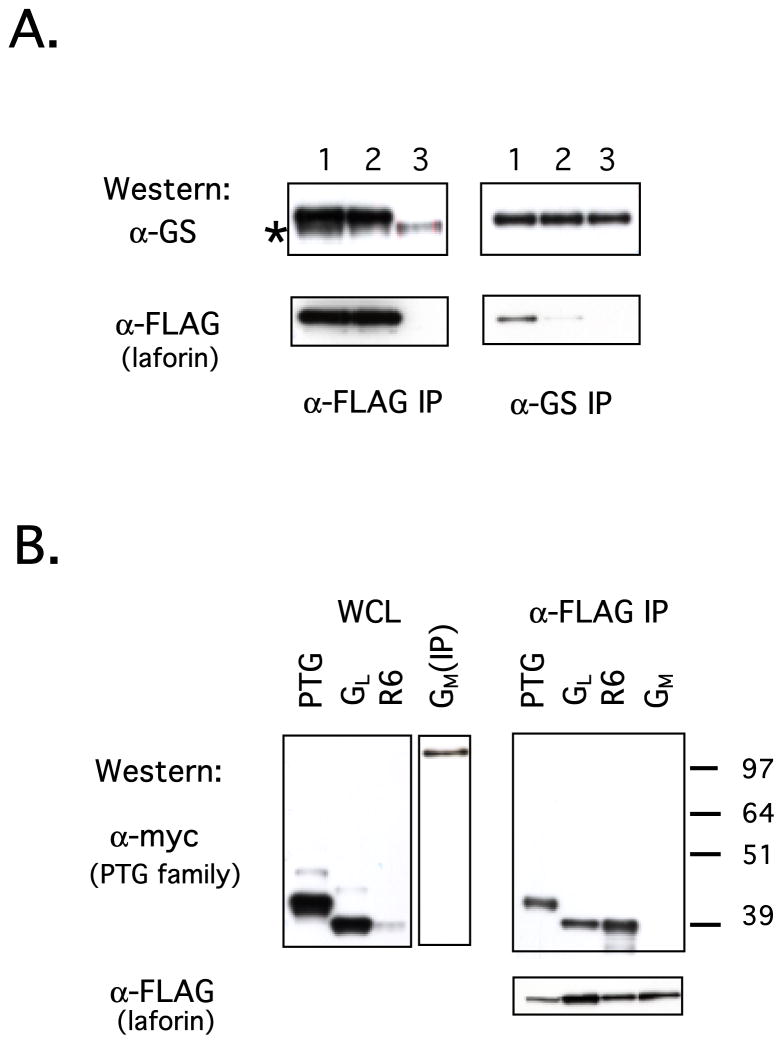
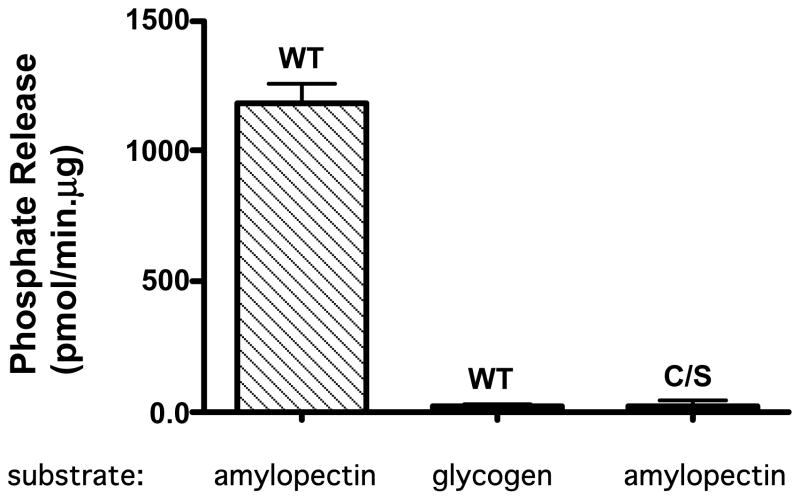
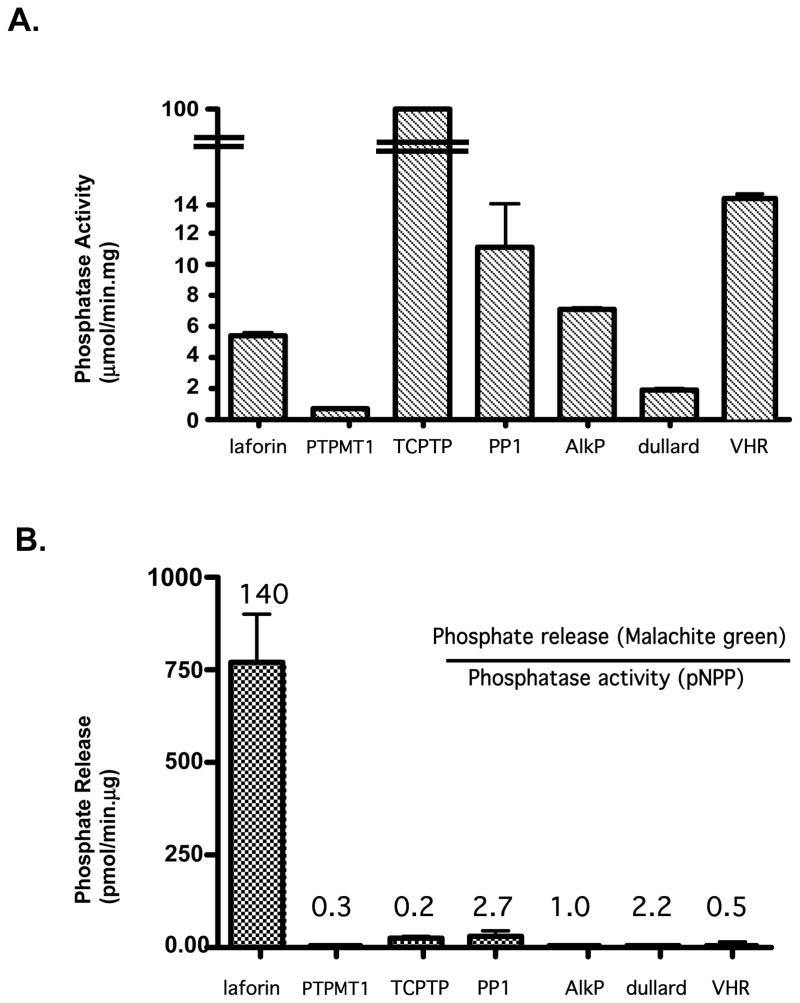
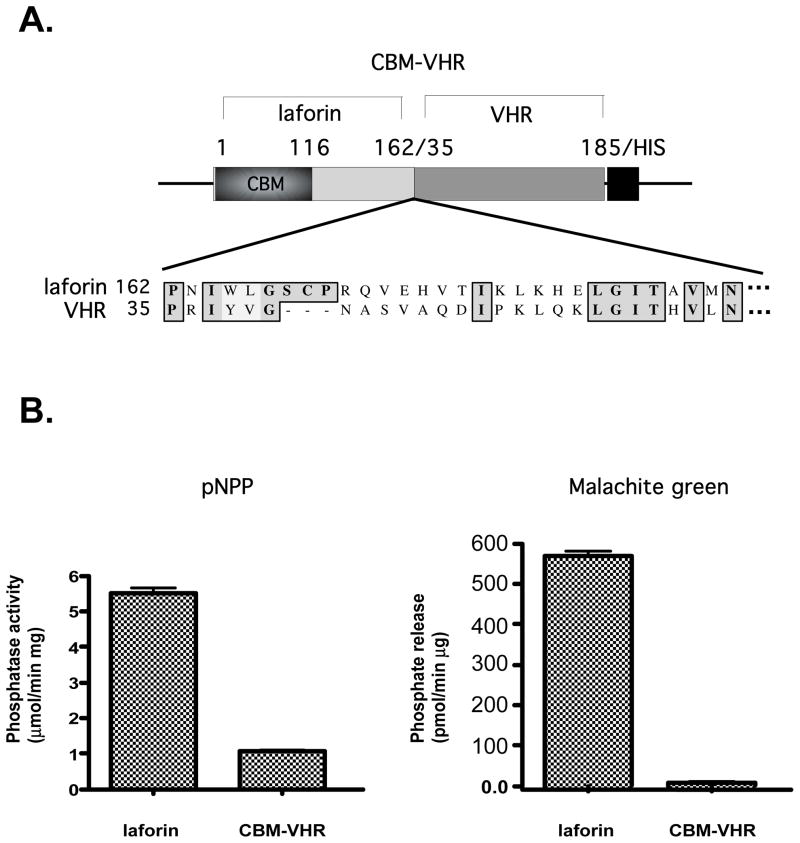
Laforin is the only phosphatase in the animal kingdom that contains a carbohydrate-binding module. Mutations in the gene encoding laforin result in Lafora disease, a fatal autosomal recessive neurodegenerative disorder, which is diagnosed by the presence of intracellular deposits of insoluble complex carbohydrates known as Lafora bodies. We demonstrate that laforin interacts with proteins known to be involved in glycogen metabolism and rule out several of these proteins as potential substrates. Surprisingly, we find that laforin displays robust phosphatase activity against a phosphorylated complex carbohydrate. Furthermore, this activity is unique to laforin, since several other phosphatases are unable to dephosphorylate polysaccharides. Finally, fusing the carbohydrate-binding module of laforin to the dual specific phosphatase VHR does not result in the ability of this phosphatase to dephosphorylate polysaccharides. Therefore, we hypothesize that laforin is unique in its ability to utilize a phosphorylated complex carbohydrate as a substrate and that this function may be necessary for the maintenance of normal cellular glycogen.
Figures
Similar articles
-
Laforin, the dual-phosphatase responsible for Lafora disease, interacts with R5 (PTG), a regulatory subunit of protein phosphatase-1 that enhances glycogen accumulation.Hum Mol Genet. 2003 Dec 1;12(23):3161-71. doi: 10.1093/hmg/ddg340. Epub 2003 Oct 7. Hum Mol Genet. 2003. PMID: 14532330
-
Expression, purification and characterization of soluble red rooster laforin as a fusion protein in Escherichia coli.BMC Biochem. 2014 Apr 2;15:8. doi: 10.1186/1471-2091-15-8. BMC Biochem. 2014. PMID: 24690255 Free PMC article.
-
Laforin, a dual specificity phosphatase involved in Lafora disease, is present mainly as monomeric form with full phosphatase activity.PLoS One. 2011;6(8):e24040. doi: 10.1371/journal.pone.0024040. Epub 2011 Aug 26. PLoS One. 2011. PMID: 21887368 Free PMC article.
-
Laforin, a protein with many faces: glucan phosphatase, adapter protein, et alii.FEBS J. 2013 Jan;280(2):525-37. doi: 10.1111/j.1742-4658.2012.08549.x. Epub 2012 Mar 16. FEBS J. 2013. PMID: 22364389 Free PMC article. Review.
-
Lafora disease: Current biology and therapeutic approaches.Rev Neurol (Paris). 2022 Apr;178(4):315-325. doi: 10.1016/j.neurol.2021.06.006. Epub 2021 Jul 21. Rev Neurol (Paris). 2022. PMID: 34301405 Free PMC article. Review.
Cited by
-
Are there errors in glycogen biosynthesis and is laforin a repair enzyme?FEBS Lett. 2011 Oct 20;585(20):3216-8. doi: 10.1016/j.febslet.2011.09.009. Epub 2011 Sep 16. FEBS Lett. 2011. PMID: 21930129 Free PMC article. Review.
-
Dimeric quaternary structure of human laforin.J Biol Chem. 2015 Feb 20;290(8):4552-4559. doi: 10.1074/jbc.M114.627406. Epub 2014 Dec 23. J Biol Chem. 2015. PMID: 25538239 Free PMC article.
-
Myofiber-type-dependent 'boulder' or 'multitudinous pebble' formations across distinct amylopectinoses.Acta Neuropathol. 2024 Feb 27;147(1):46. doi: 10.1007/s00401-024-02698-x. Acta Neuropathol. 2024. PMID: 38411740
-
Trehalose Ameliorates Seizure Susceptibility in Lafora Disease Mouse Models by Suppressing Neuroinflammation and Endoplasmic Reticulum Stress.Mol Neurobiol. 2021 Mar;58(3):1088-1101. doi: 10.1007/s12035-020-02170-3. Epub 2020 Oct 22. Mol Neurobiol. 2021. PMID: 33094475
-
Deciphering the role of malin in the lafora progressive myoclonus epilepsy.IUBMB Life. 2012 Oct;64(10):801-8. doi: 10.1002/iub.1072. Epub 2012 Jul 20. IUBMB Life. 2012. PMID: 22815132 Free PMC article. Review.
References
-
- Berkovic SF, Andermann F, Carpenter S, Wolfe LS. N Engl J Med. 1986;315(5):296–305. - PubMed
-
- Lafora G. Z Ges Neurol Psychiatr. 1911;6:1–14.
-
- Minassian BA. Adv Neurol. 2002;89:199–210. - PubMed
-
- Lehesjoki AE. Adv Neurol. 2002;89:193–197. - PubMed
-
- Minassian BA, Lee JR, Herbrick JA, Huizenga J, Soder S, Mungall AJ, Dunham I, Gardner R, Fong CY, Carpenter S, Jardim L, Satishchandra P, Andermann E, Snead OC, 3rd, Lopes-Cendes I, Tsui LC, Delgado-Escueta AV, Rouleau GA, Scherer SW. Nat Genet. 1998;20(2):171–174. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
