

A transgenic approach to understanding the influence of carbonic anhydrase on C18OO discrimination during C4 photosynthesis
- PMID: 16905667
- PMCID: PMC1586065
- DOI: 10.1104/pp.106.085167
A transgenic approach to understanding the influence of carbonic anhydrase on C18OO discrimination during C4 photosynthesis
Abstract
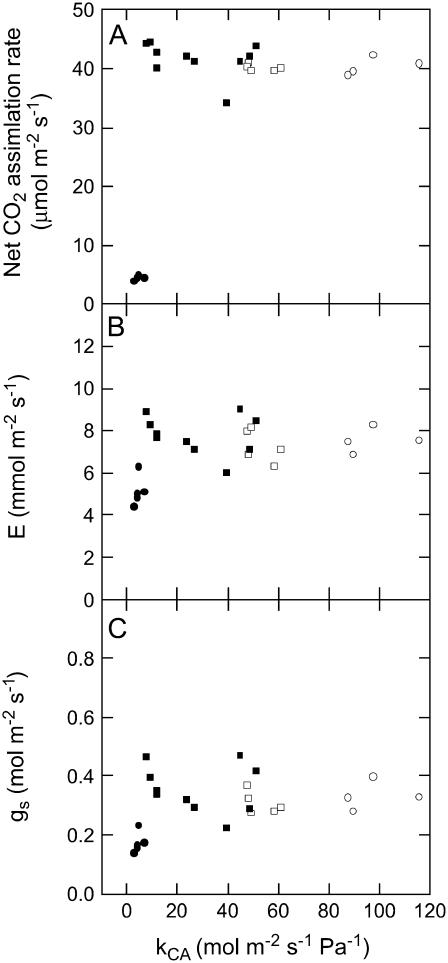
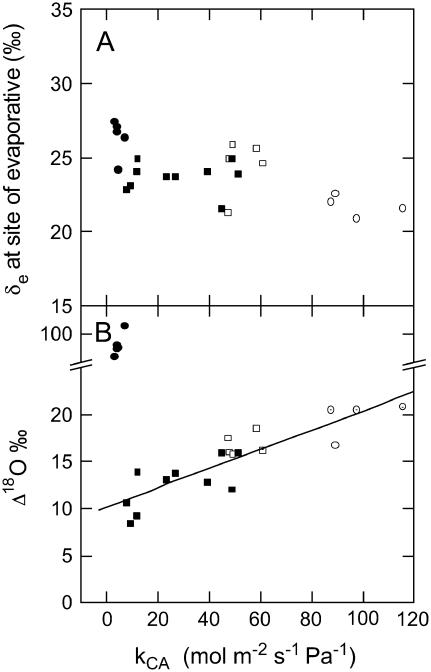
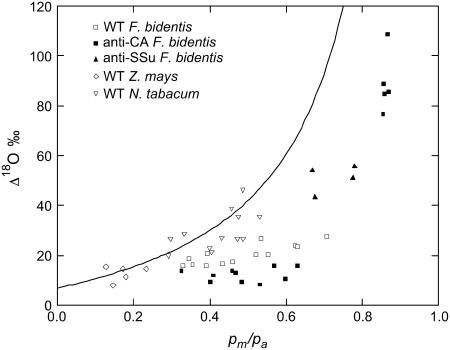
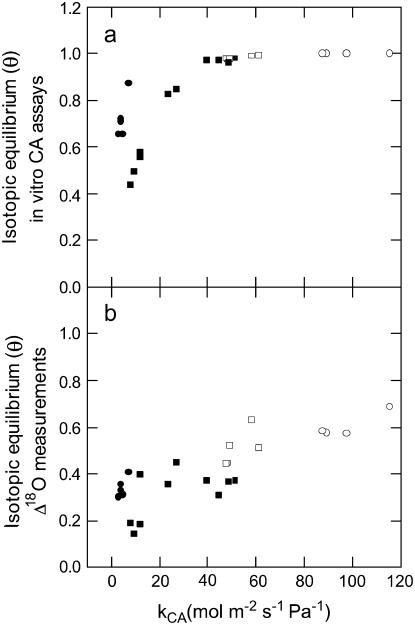
The oxygen isotope composition of atmospheric CO(2) is an important signal that helps distinguish between ecosystem photosynthetic and respiratory processes. In C(4) plants the carbonic anhydrase (CA)-catalyzed interconversion of CO(2) and bicarbonate (HCO(3)(-)) is an essential first reaction for C(4) photosynthesis but also plays an important role in the CO(2)-H(2)O exchange of oxygen as it enhances the rate of isotopic equilibrium between CO(2) and water. The C(4) dicot Flaveria bidentis containing genetically reduced levels of leaf CA (CA(leaf)) has been used to test whether changing leaf CA activity influences online measurements of C(18)OO discrimination (Delta(18)O) and the proportion of CO(2) in isotopic equilibrium with leaf water at the site of oxygen exchange (theta). The Delta(18)O in wild-type F. bidentis, which contains high levels of CA relative to the rates of net CO(2) assimilation, was less than predicted by models of Delta(18)O. Additionally, Delta(18)O was sensitive to small decreases in CA(leaf). However, reduced CA activity in F. bidentis had little effect on net CO(2) assimilation, transpiration rates (E), and stomatal conductance (g(s)) until CA levels were less than 20% of wild type. The values of theta determined from measurements of Delta(18)O and the (18)O isotopic composition of leaf water at the site of evaporation (delta(e)) were low in the wild-type F. bidentis and decreased in transgenic plants with reduced levels of CA activity. Measured values of theta were always significantly lower than the values of theta predicted from in vitro CA activity and gas exchange. The data presented here indicates that CA content in a C(4) leaf may not represent the CA activity associated with the CO(2)-H(2)O oxygen exchange and therefore may not be a good predictor of theta during C(4) photosynthesis. Furthermore, uncertainties in the isotopic composition of water at the site of exchange may also limit the ability to accurately predict theta in C(4) plants.
Figures
References
-
- Affek HP, Krisch MJ, Yakir D (2006) Effects of intraleaf variations in carbonic anhydrase activity and gas exchange on leaf (COO)-O-18 isoflux in Zea mays. New Phytol 169: 321–329 - PubMed
-
- Badger M (2003) The roles of carbonic anhydrases in photosynthetic CO2 concentrating mechanism. Photosynth Res 77: 83–94 - PubMed
-
- Badger M, Price GD (1994) The role of carbonic anhydrase in photosynthesis. Annu Rev Plant Physiol Plant Mol Biol 45: 369–392
-
- Barbour MM, Farquhar GD (2004) Do pathways of water movement and leaf anatomical dimensions allow development of gradients in (H2O)-O-18 between veins and the sites of evaporation within leaves? Plant Cell Environ 27: 107–121
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
