

Biosynthesis of Dictyostelium discoideum differentiation-inducing factor by a hybrid type I fatty acid-type III polyketide synthase
- PMID: 16906151
- PMCID: PMC2864586
- DOI: 10.1038/nchembio811
Biosynthesis of Dictyostelium discoideum differentiation-inducing factor by a hybrid type I fatty acid-type III polyketide synthase
Abstract
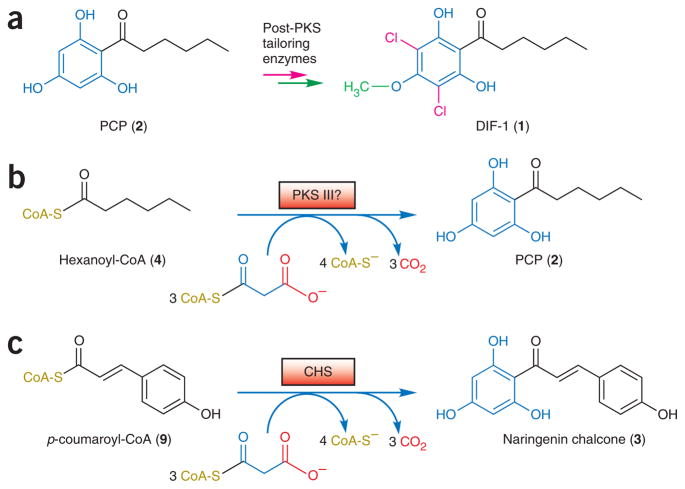
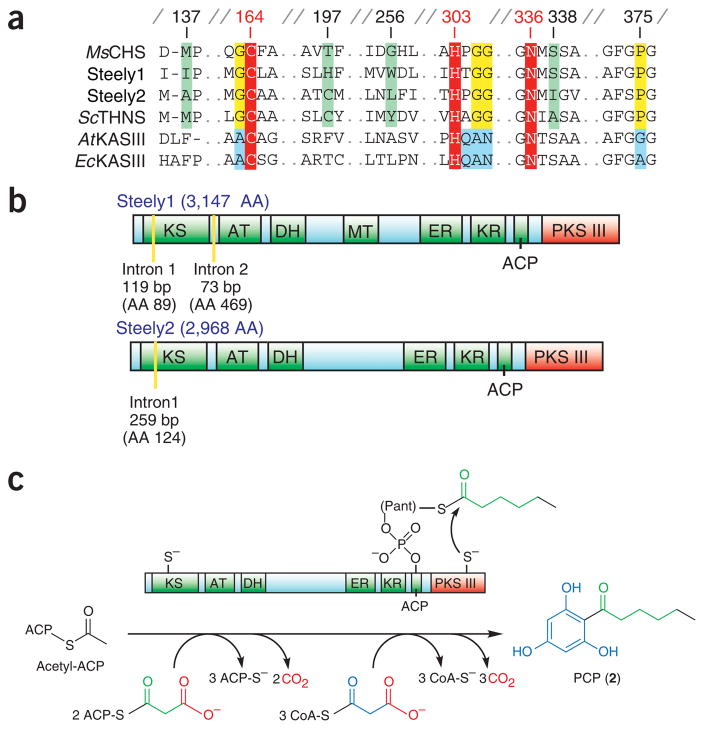
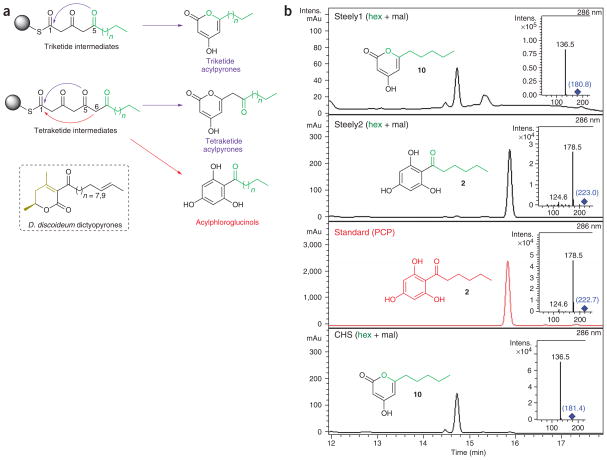
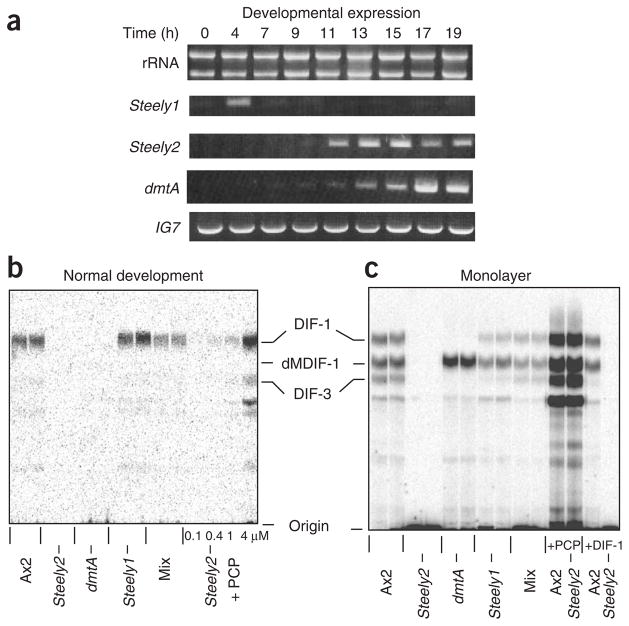
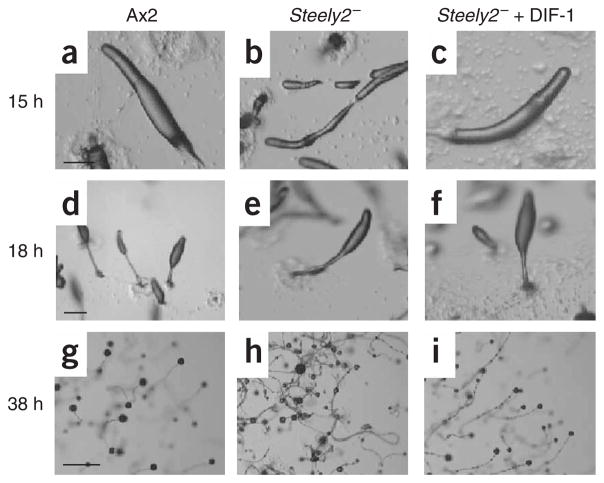
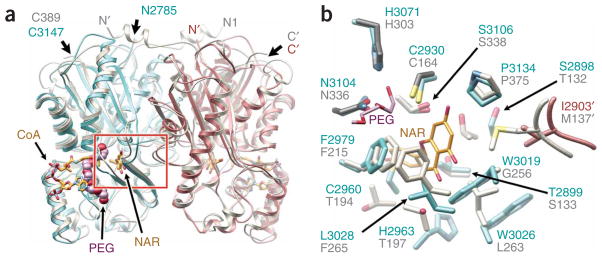
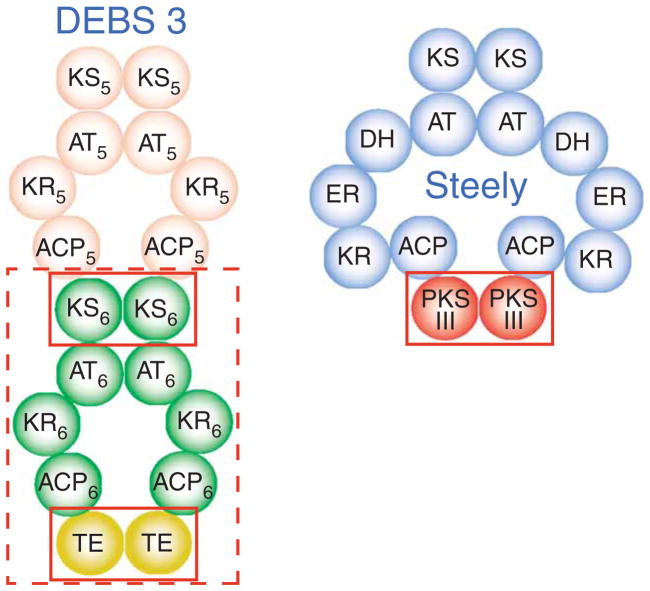
Differentiation-inducing factors (DIFs) are well known to modulate formation of distinct communal cell types from identical Dictyostelium discoideum amoebas, but DIF biosynthesis remains obscure. We report complimentary in vivo and in vitro experiments identifying one of two approximately 3,000-residue D. discoideum proteins, termed 'steely', as responsible for biosynthesis of the DIF acylphloroglucinol scaffold. Steely proteins possess six catalytic domains homologous to metazoan type I fatty acid synthases (FASs) but feature an iterative type III polyketide synthase (PKS) in place of the expected FAS C-terminal thioesterase used to off load fatty acid products. This new domain arrangement likely facilitates covalent transfer of steely N-terminal acyl products directly to the C-terminal type III PKS active sites, which catalyze both iterative polyketide extension and cyclization. The crystal structure of a steely C-terminal domain confirms conservation of the homodimeric type III PKS fold. These findings suggest new bioengineering strategies for expanding the scope of fatty acid and polyketide biosynthesis.
Conflict of interest statement
The authors declare that they have no competing financial interests.
Figures
Comment in
-
A type III PKS makes the DIFference.Nat Chem Biol. 2006 Sep;2(9):451-2. doi: 10.1038/nchembio0906-451. Nat Chem Biol. 2006. PMID: 16921353 No abstract available.
References
-
- Kessin RH. Dictyostelium. Cambridge University Press; Cambridge, UK: 2001.
-
- Morris HR, Taylor GW, Masento MS, Jermyn KA, Kay RR. Chemical structure of the morphogen differentiation inducing factor from Dictyostelium discoideum. Nature. 1987;328:811–814. - PubMed
-
- Thompson CR, Kay RR. The role of DIF-1 signaling in Dictyostelium development. Mol Cell. 2000;6:1509–1514. - PubMed
-
- Gokan N, et al. Structural requirements of Dictyostelium differentiation-inducing factors for their stalk-cell-inducing activity in Dictyostelium cells and anti-proliferative activity in K562 human leukemic cells. Biochem Pharmacol. 2005;70:676–685. - PubMed
-
- Kay RR. The biosynthesis of differentiation-inducing factor, a chlorinated signal molecule regulating Dictyostelium development. J Biol Chem. 1998;273:2669–2675. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- PubChem-Substance/12011237
- PubChem-Substance/12011238
- PubChem-Substance/12011239
- PubChem-Substance/12011240
- PubChem-Substance/12011241
- PubChem-Substance/12011242
- PubChem-Substance/12011243
- PubChem-Substance/12011244
- PubChem-Substance/12011245
- PubChem-Substance/12011246
- PubChem-Substance/12011247
- PubChem-Substance/12011248
- PubChem-Substance/12011249
- PubChem-Substance/12011250
- PubChem-Substance/12011251
- PubChem-Substance/12011252
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
