

Human T-cell leukemia virus type 1 (HTLV-1) and HTLV-2 use different receptor complexes to enter T cells
- PMID: 16912281
- PMCID: PMC1563841
- DOI: 10.1128/JVI.00389-06
Human T-cell leukemia virus type 1 (HTLV-1) and HTLV-2 use different receptor complexes to enter T cells
Abstract
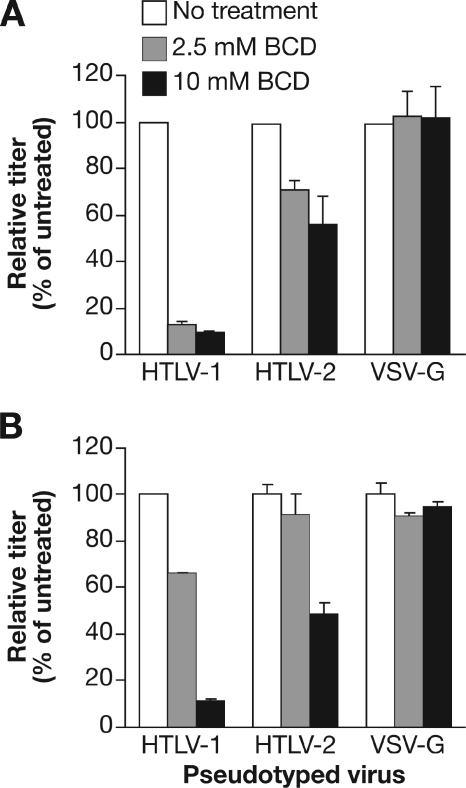
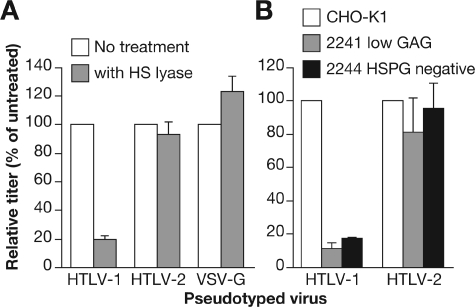
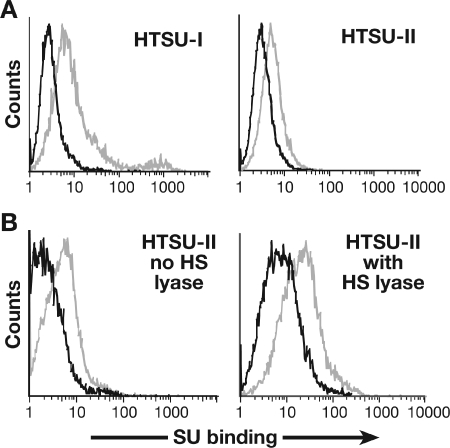
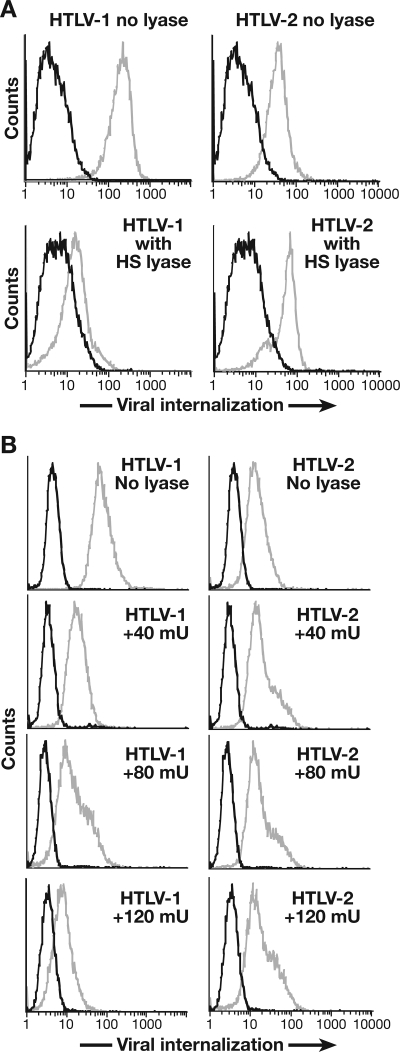
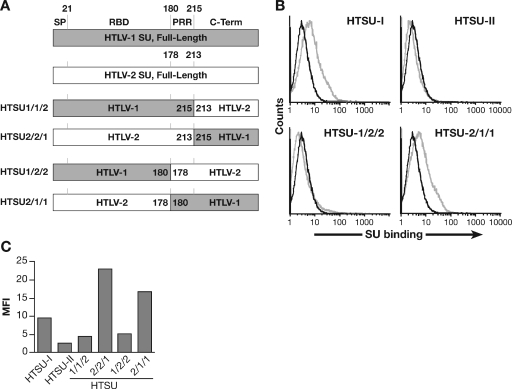
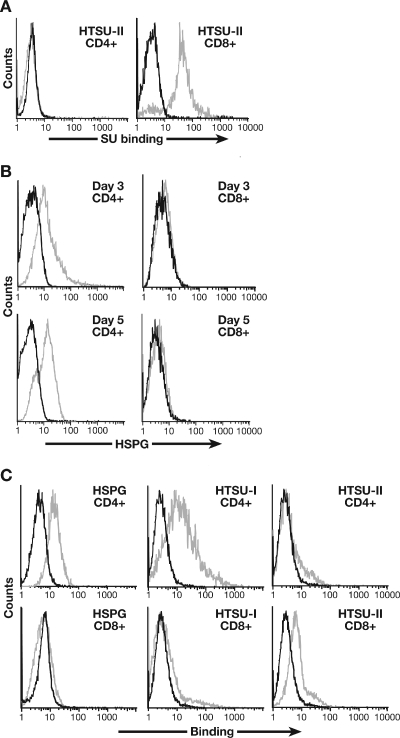
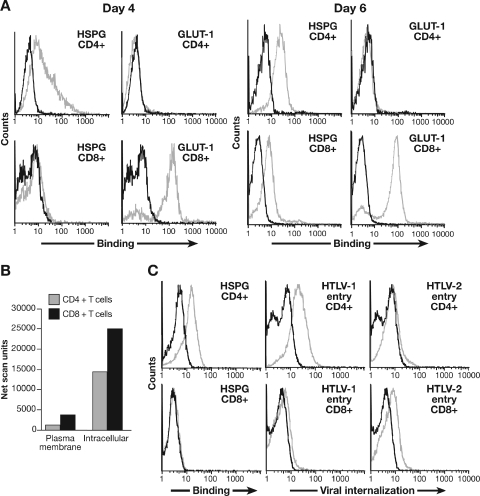
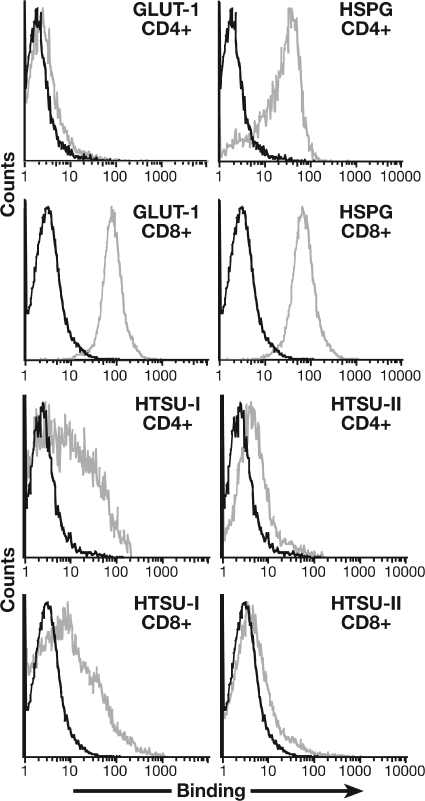
Studies using adherent cell lines have shown that glucose transporter-1 (GLUT-1) can function as a receptor for human T-cell leukemia virus type 1 (HTLV). In primary CD4(+) T cells, heparan sulfate proteoglycans (HSPGs) are required for efficient entry of HTLV-1. Here, the roles of HSPGs and GLUT-1 in HTLV-1 and HTLV-2 Env-mediated binding and entry into primary T cells were studied. Examination of the cell surface of activated primary T cells revealed that CD4(+) T cells, the primary target of HTLV-1, expressed significantly higher levels of HSPGs than CD8(+) T cells. Conversely, CD8(+) T cells, the primary target of HTLV-2, expressed GLUT-1 at dramatically higher levels than CD4(+) T cells. Under these conditions, the HTLV-2 surface glycoprotein (SU) binding and viral entry were markedly higher on CD8(+) T cells while HTLV-1 SU binding and viral entry were higher on CD4(+) T cells. Binding studies with HTLV-1/HTLV-2 SU recombinants showed that preferential binding to CD4(+) T cells expressing high levels of HSPGs mapped to the C-terminal portion of SU. Transfection studies revealed that overexpression of GLUT-1 in CD4(+) T cells increased HTLV-2 entry, while expression of HSPGs on CD8(+) T cells increased entry of HTLV-1. These studies demonstrate that HTLV-1 and HTLV-2 differ in their T-cell entry requirements and suggest that the differences in the in vitro cellular tropism for transformation and in vivo pathobiology of these viruses reflect different interactions between their Env proteins and molecules on CD4(+) and CD8(+) T cells involved in entry.
Figures

References
-
- Albritton, L. M., L. Tseng, D. Scadden, and J. M. Cunningham. 1989. A putative murine ecotropic retrovirus receptor gene encodes a multiple membrane-spanning protein and confers susceptibility to virus infection. Cell 57:659-666. - PubMed
-
- Al-Hasani, H., D. R. Yver, and S. W. Cushman. 1999. Overexpression of the glucose transporter GLUT4 in adipose cells interferes with insulin-stimulated translocation. FEBS Lett. 460:338-342. - PubMed
-
- Argyris, E. G., E. Acheampong, G. Nunnari, M. Mukhtar, K. J. Williams, and R. J. Pomerantz. 2003. Human immunodeficiency virus type 1 enters primary human brain microvascular endothelial cells by a mechanism involving cell surface proteoglycans independent of lipid rafts. J. Virol. 77:12140-12151. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
