

Potential role for p53 in the permissive life cycle of human cytomegalovirus
- PMID: 16912290
- PMCID: PMC1563868
- DOI: 10.1128/JVI.00505-06
Potential role for p53 in the permissive life cycle of human cytomegalovirus
Abstract
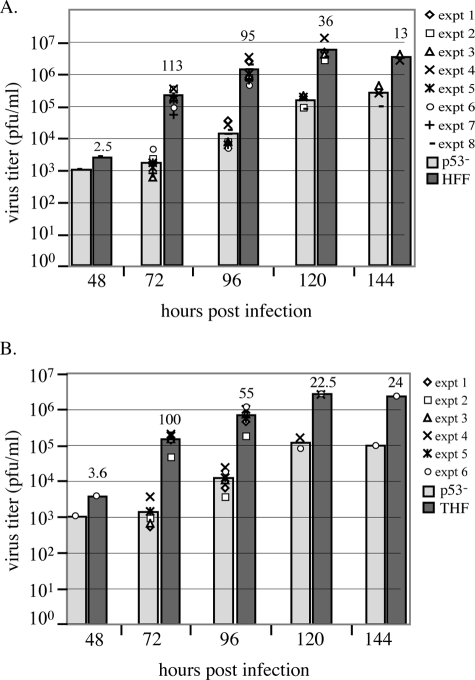
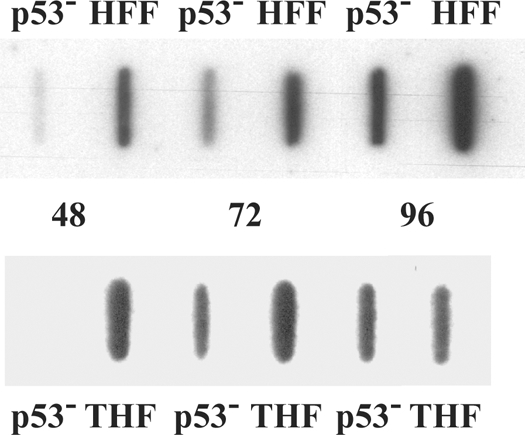
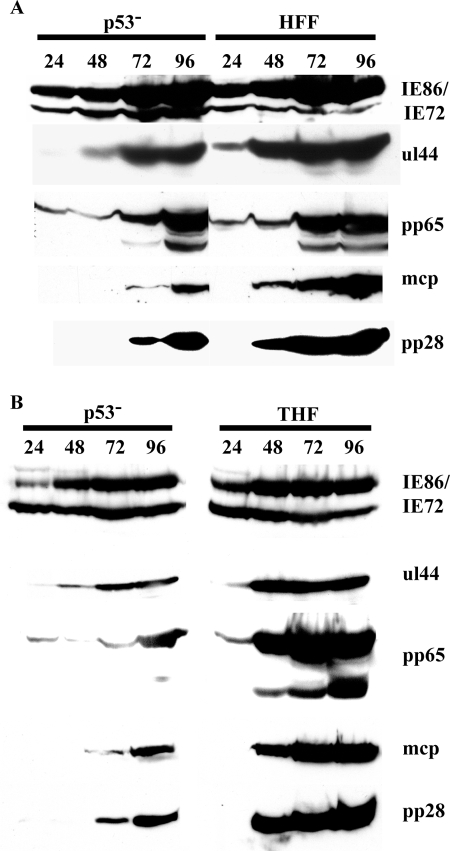
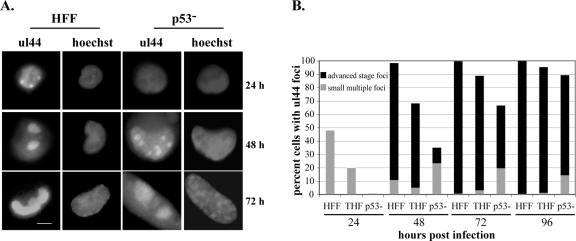
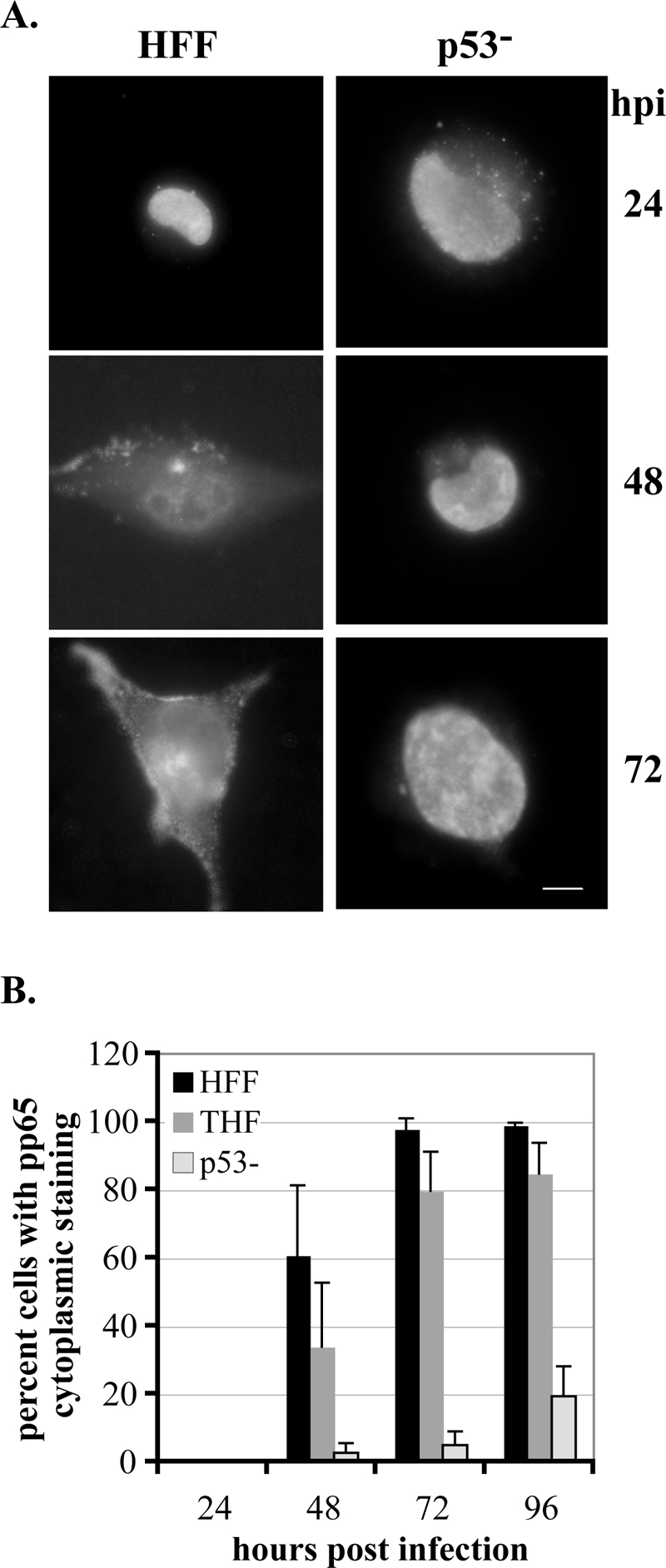
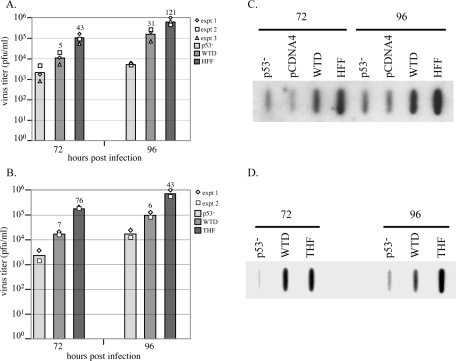
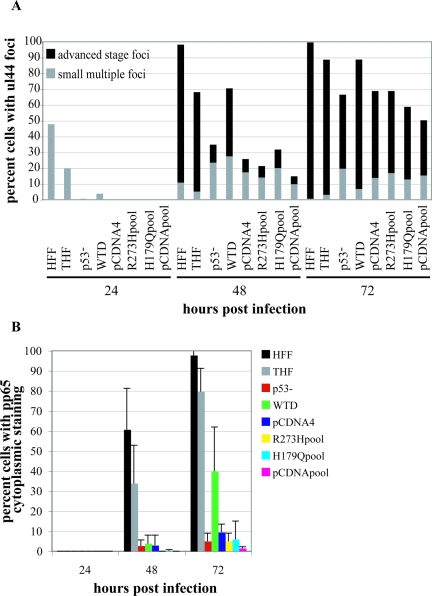
Infection of primary fibroblasts with human cytomegalovirus (HCMV) causes a rapid stabilization of the cellular protein p53. p53 is a major effector of the cellular damage response, and activation of this transcription factor can lead either to cell cycle arrest or to apoptosis. Viruses employ many tactics to avoid p53-mediated effects. One method HCMV uses to counteract p53 is sequestration into its viral replication centers. In order to determine whether or not HCMV benefits from this sequestration, we infected a p53(-/-) fibroblast line. We find that although these cells are permissive for viral infection, several parameters are substantially altered compared to wild-type (wt) fibroblasts. p53(-/-) cells show delayed and decreased accumulation of infectious viral particles compared to control fibroblasts, with the largest difference of 100-fold at 72 h post infection (p.i.) and peak titers decreased by approximately 10- to 20-fold at 144 h p.i. Viral DNA accumulation is also delayed and somewhat decreased in p53(-/-) cells; however, on average, levels of DNA are not more than fivefold lower than wt at any time p.i. and thus cannot account entirely for the observed differences in titers. In addition, there are delays in the expression of several key viral proteins, including the early replication protein UL44 and some of the late structural proteins, pp28 (UL99) and MCP (UL86). UL44 localization also indicates delayed formation and maturation of the replication centers throughout the course of infection. Localization of the major tegument protein pp65 (UL83) is also altered in these p53(-/-) cells. Partial reconstitution of the p53(-/-) cells with a wt copy of p53 returns all parameters toward wt, while reconstitution with mutant p53 does not. Taken together, our data suggest that wt p53 enhances the ability of HCMV to replicate and produce high concentrations of infectious virions in permissive cells.
Figures
References
-
- Albrecht, T., M. P. Fons, I. Bologh, S. AbuBakar, C. Z. Deng, and D. Millinoff. 1991. Metabolic and cellular effects of human cytomegalovirus infection. Transplant. Proc. 23:48-55. - PubMed
-
- Boldogh, I., S. AbuBakar, and T. Albrecht. 1990. Activation of proto-oncogenes: an immediate early event in human cytomegalovirus infection. Science 247:561-564. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
