

Elongation complexes of Thermus thermophilus RNA polymerase that possess distinct translocation conformations
- PMID: 16914440
- PMCID: PMC1557819
- DOI: 10.1093/nar/gkl559
Elongation complexes of Thermus thermophilus RNA polymerase that possess distinct translocation conformations
Abstract
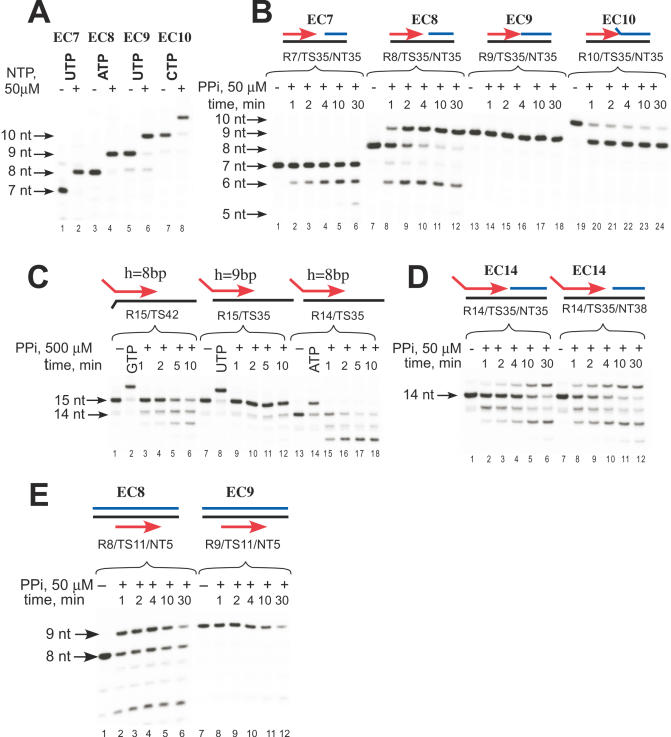
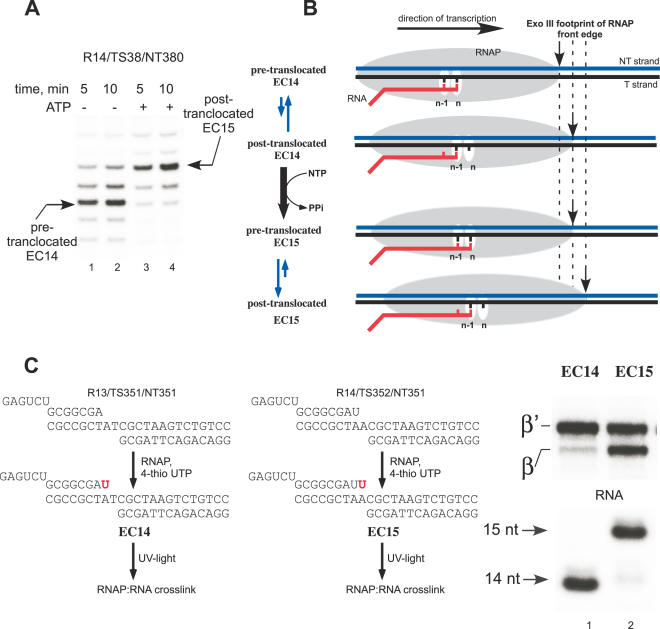
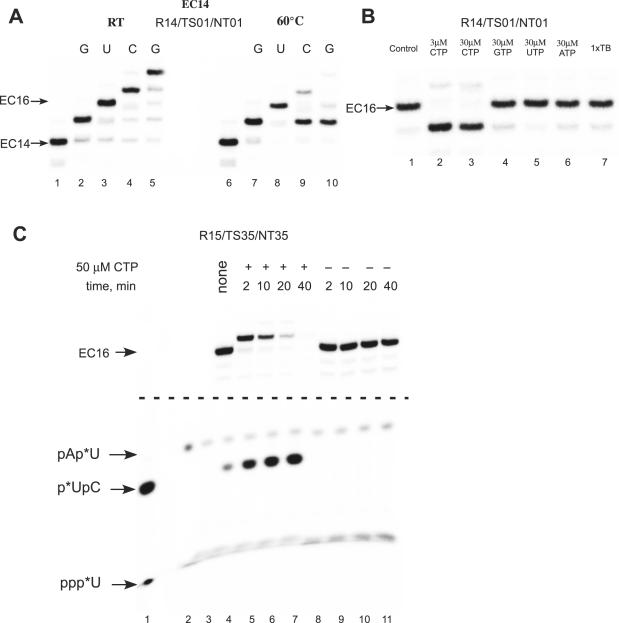
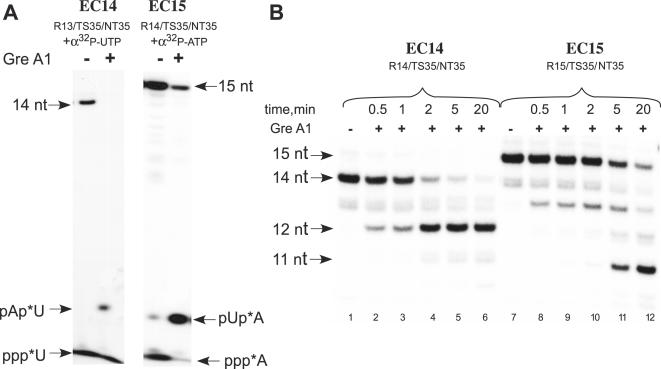
We have characterized elongation complexes (ECs) of RNA polymerase from the extremely thermophilic bacterium, Thermus thermophilus. We found that complexes assembled on nucleic acid scaffolds are transcriptionally competent at high temperature (50-80 degrees C) and, depending upon the organization of the scaffold, possess distinct translocation conformations. ECs assembled on scaffolds with a 9 bp RNA:DNA hybrid are highly stable, resistant to pyrophosphorolysis, and are in the posttranslocated state. ECs with an RNA:DNA hybrid longer or shorter than 9 bp appear to be in a pretranslocated state, as evidenced by their sensitivity to pyrophosphorolysis, GreA-induced cleavage, and exonuclease footprinting. Both pretranslocated (8 bp RNA:DNA hybrid) and posttranslocated (9 bp RNA:DNA hybrid) complexes were crystallized in distinct crystal forms, supporting the homogeneity of the conformational states in these complexes. Crystals of a posttranslocated complex were used to collect diffraction data at atomic resolution.
Figures

References
-
- Campbell E.A., Korzheva N., Mustaev A., Murakami K., Nair S., Goldfarb A., Darst S.A. Structural mechanism for rifampicin inhibition of bacterial RNA polymerase. Cell. 2001;104:901–912. - PubMed
-
- Murakami K.S., Masuda S., Darst S.A. Structural basis of transcription initiation: RNA polymerase holoenzyme at 4 A resolution. Science. 2002;296:1280–1284. - PubMed
-
- Zhang G., Campbell E.A., Minakhin L., Richter C., Severinov K., Darst S.A. Crystal structure of Thermus aquaticus core RNA polymerase at 3.3 A resolution. Cell. 1999;98:811–824. - PubMed
-
- Artsimovitch I., Patlan V., Sekine S., Vassylyeva M.N., Hosaka T., Ochi K., Yokoyama S., Vassylyev D.G. Structural basis for transcription regulation by alarmone ppGpp. Cell. 2004;117:299–310. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
